Introduction
Auricularia species have ear-shaped fruiting bodies and are considered a wood-decaying fungus which commonly grows on wood. They are easily cultivated, and some species have high economic, nutritional, medicinal, and pharmacological values in Asian countries [1,2]. While the Index Fungorum (http://www.indexfungorum.org/) lists 177 species and variety names for Auricularia, only 10 to 15 identified species have been studied [3]. Auricularia species are well-known for their morphological characteristics and distribution, but species level identification can be difficult due to their similar colors, fruit bodies, and sizes. Auricularia mesenterica is a type species of Auricularia [4]. A recent study [5] showed that A. auricula-judae has been treated phylogenetically as a species complex. Some specimens considered to be A. auricula-judae were later identified as A. heimuer, A. minutissima, A. thailandica, and A. villosula [6,7].
Since 1978, A. hispida Iwade, A. mesenterica (Dicks.) Pers., and A. polytricha (Mont.) Sacc. (current name: A. nigricans (Sw.) Birkebak, Looney, and Sánchez-García) have been recorded in Korea [8,9]. Recently, two unrecorded species (Auricularia cornea and A. minutissima) were reported based on morphological and molecular phylogenetic analysis in Korea [10], but comprehensive phylogenic and taxonomic studies for the identification of Auricularia in Korea have not yet been conducted. Based on DNA sequence data comparisons and morphological characteristics from 127 Auricularia specimens deposited in the Korea National Arboretum (KH), Korea five species of Auricularia were identified, of which A. americana, A. heimuer, and A. villosula were previously unidentified in Korea. Moreover, phylogenetic analysis demonstrated that A. auricula-judae, which had previously been identified in Korea, was not present in these samples.
Materials and Methods
Fungal materials and morphological observations
A total of 127 Auricularia samples were collected from 2008 to 2020 from various urban parks, legally protected trees, and forests in Korea. Forest floors, leaf litter, twigs, fallen trunks, leaves, and soil were inspected for collections. Macro-morphological characteristics were examined using a dissecting microscope. For micro-morphological characterization, abhymenial hairs, basidiospores, and basidia were mounted in distilled water and 5% KOH and examined using a compound microscope (Olympus BX53, Tokyo, Japan) and photographed with a Jenoptik ProgRes C14 Plus Camera (Jenoptik Corporation, Jena, Germany). Microscopic parameters were measured and calculated using the ProgRes Capture Pro v.2.8.8. software (Jenoptik Corporation, Jena, Germany). Dried specimens were deposited in the herbarium of the Korea National Arboretum (KH).
DNA isolation, amplification, sequencing, and phylogenetic analysis
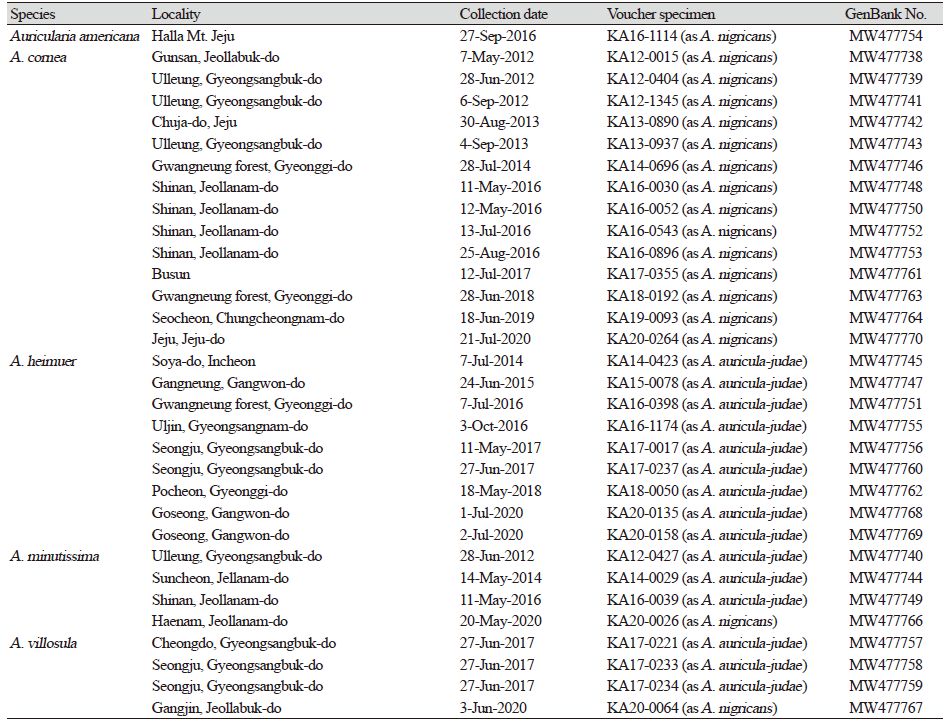
Genomic DNA from specimens was extracted using the DNeasy Plant Mini DNA Extraction Kit (Qiagen Inc., Valencia, CA, USA), according to the manufacturer’s instructions. After dividing into morphology groups, a representative sample was selected from each group. Of the 127 specimens, 33 samples were selected as a representative sample and these samples were successfully extracted in this study. The extracted DNA was used for PCR amplification. The internal transcribed spacer (ITS) was amplified with primers ITS5 and ITS4 [11]. The PCR amplicons were purified using a QIAquick purification Kit (Qiagen Inc., Hilden, Germany), and then directly sequenced using an ABI Prism® 377 Automatic DNA Sequencer (Applied Biosystems, Foster City, CA, USA) with a BigDye Cycle Sequencing Kit Version 3.1 (Applied Biosystems). A BLASTN search against the NCBI database (http://www.ncbi.nlm.nih.gov) was carried out, and new sequences were deposited in GenBank (www.ncbi.nlm.nih.gov/genbank/). Sequences of Auricularia used in the phylogenetic tree were downloaded from GenBank, and Exidia grandulosa (AF291279) was used as an outgroup. For phylogenetic analyses, the data sets were aligned in MAFFT version 7.475 [12], using the auto option. The DNA sequences generated in this study were assembled and edited using BioEdit v. 7.2.5 [13] and MEGA v. 7.0.26 [14]. A phylogenetic tree was constructed using RAxML in the CIPRES Science Gateway (https://www.phylo.org). The relative robustness of the individual branches was estimated by bootstrapping using 1,000 replicates. The GenBank accession numbers of the ITS sequences used in this study are listed in Table 1.
Results
Phylogenetic analysis
A total of 33 ITS sequences were obtained from the Auricularia specimens in this study (Table 1). Three unrecorded species (A. americana, A. heimuer and A. villosula) were resolved by conducting RAxML analysis of the ITS sequences. Although there was sequence variation, they formed well-supported clades. A previous study [10] showed that phylogenetic position of A. cornea and A. minutissima with one sample, respectively. In this study, A. cornea and A. minutissima form a distinct clade with additional samples from Korea National Arboretum. A. minutissma formed a sister group to the A. heimuer in ML analysis. A previous study demonstrated A. auricula-judae forming a species complex with ITS sequence data (not included Korean samples here) [5]. In this study, the Korean samples considered to be A. auricula-judae before were identified as A. heimuer, A. minutissima, and A. villosula. Therefore, the phylogenetic positions of those species were confirmed in this study.
Taxonomy
Auricularia americana Parmasto & I. Parmasto ex Audet, Boulet & Sirard, in Boulet, Les Champignons des Arbres de l'Est de l'Amérique du Nord: 287. 2003. (Figs. 1 and 2)
Korean name: Jeonnamu-Mogi (전나무목이); derived from the Korean family name of host (Jeonnamu), ‘Abies’
Description: Basidiocarps solitary or gregarious, sessile or stipitate, auriculate to cupulate, with undulate margin, hard-gelatinous to cartilaginous, 2-4 cm broad, 0.4-1 mm thick; upper surface velvety with folds, olive brown to light brown; lower surface smooth with folds, light brown. Medulla absent or indistinct. Internal hyphae branched, septate, thin-walled, hyaline, with clamp connections, 1.5-3 μm, diam. Abhymenial hairs solitary or gregarious, cylindrical, flexuous, thick-walled with a narrow lumen, apically acute to obtuse, hyaline with pale brown pigmentation at the base, 45-153×7-11 μm. Basidia narrowly clavate to cylindrical, transversely 3-septate, hyaline, 60-75×4-6 μm. Basidiospores allantoid to reniform, thin-walled, smooth, hyaline, 11-14×4-5.5 μm.
Notes: The Abies balsamea (L.) Mill. was recorded as a host of the holotype of A. americana (Index Fungorum; www.indexfungorum.org/). The Korean specimen (KA16-1114) was collected from Abies koreana E.H. Wilson on Jeju Island. According to a previous report [4], this species grows exclusively on gymnosperms. To date, this Korean collection has been preserved in KH.
Auricularia heimuer F. Wu, B.K. Cui & Y.C. Dai, in Wu, Yuan, Malysheva, Du & Dai, Phytotaxa 186: 248. (Figs. 1 and 3)
Korean name: Heung-Mogi (흑목이); as ‘heimuer’is a black jelly fungi in Chinese; ‘hei’ means black and ‘muer’ means jelly fungi.
Description: Basidiocarps solitary or gregarious, sessile or stipitate, auriculate to cupulate, with undulate margin, hard-gelatinous to cartilaginous, 2-5 cm broad, 0.5-1 mm thick; upper surface velvety with folds, olive brown to light brown; lower surface smooth with folds, light brown. Medulla pale brown, 115-180 μm, broad. Internal hyphae branched, septate, thin-walled, hyaline, with clamp connections, 2-4 μm, diam. Abhymenial hairs are solitary to gregarious, cylindrical, flexuous, thick-walled with a narrow lumen, apically acute to obtuse, hyaline with pale brown pigmentation at the base, 45-140×4.5-7 μm. Basidia narrowly clavate to cylindrical, transversely 3-septate, hyaline, 35-58×4.5-5.5 μm. Basidiospores allantoid, reniform, thin-walled, hyaline, 9.5-14×5-6 μm.

Notes: This species is the third most important cultivated edible mushroom worldwide [4,5]. Several studies of this species have recently been conducted using high-throughput sequencing technologies at the genomic level [15,16]. In Korea, specimens of A. heimuer were considered to be part of the A. auricula-judae complex. Morphological examination showed that A. heimuer produces smaller basidia (35-58 μm long vs. 64-85 μm long) and basidiospores (9.5-14 μm long vs. 14.7-18.2 μm long) than A. auricula-judae [4]. In addition, these two species are phylogenetically distant. Cultivated samples considered to be A. auricula-judae from Korea were morphologically and phylogenetically studied in this investigation. As a result of the phylogentic analysis, these samples were identified as A. heimuer, and A. auricula-judae was not identified.
Auricularia villosula Malysheva, Malysheva & Bulakh, Nov. sist. Niz. Rast. 48: 174. 2014. (Figs. 1 and 4)
Korean name: Jjalpeunteol-Mogi (짧은털목이); derived from the species ephithet ‘villosula’, Latin meaning slightly hairy
Description: Basidiocarps solitary or gregarious, sessile or stipitate, auriculate to cupulate, with undulate margin, hard-gelatinous to cartilaginous, 1-7 cm broad, 1 mm thick; upper surface velvety with folds, brown to light brown; lower surface smooth with folds, light brown. Medulla absent or indistinct. Internal hyphae branched, septate, thin-walled, hyaline, with clamp connections, 2-4.5 μm, diam. Abhymenial hairs solitary or gregarious, cylindrical, flexuous, thick-walled with a narrow lumen, apically acute to obtuse, hyaline with pale brown pigmentation at the base, 30-93×5.5-6 μm. Basidia narrowly clavate to cylindrical, transversely 3-septate, hyaline, 52-62×4.5-6.5 μm. Basidiospores allantoid to reniform, thin-walled, hyaline, 15-17×5.5-7 μm.
Notes: These specimens were also considered as A. auricula-judae complexes. Morphological comparisons showed that this species has shorter abhymenial hairs (30-93 μm long vs. 100-150 μm long) and basidia (52-62 μm long vs. 64-85 μm long) than A. auricula-judae [4]. Furthermore, phylogenetic analysis showed that A. villosula is distinctly different from A. auricula-judae.

Key to Auricularia species in Korea
Based on the collected specimens, we have provided a morphological distinguishing key for the five Auricularia species identified, as follows:
1. Abhymenial hairs <100 μm long·····················································································2
1. Abhymenial hairs >100 μm long·····················································································3
2. Basidiospores usually >15 μm long································································A. minutissima
2. Basidiospores usually <15 μm long·····································································A. villosula
3. Basidia >60 μm long·······················································································A. heimuer
3. Basidia <60 μm long···································································································4
4. Medulla present·······························································································A. cornea
4. Medulla absent or indistinct·············································································A. americana
Discussion
Phylogenetic analysis confirmed five different species in this study. Of those, three species (A. americana, A. heimuer and A. villosula) were new to Korea, while A. cornea and A. minutissima have recently been described [10].
To date, six species of Auricularia have been recorded in Korea. Auricularia auricula-judae (Bull.) Quél. is a well-known fungus as judas ear mushroom in Auriculariales. In Korea, this species was recorded on Ulleung Island in 1959 for the first time. A. auricula-judae had previously been reported in Korea [17-20], but its taxonomical data had not been investigated. In Korea, phylogenetic studies of A. auricula-judae have never been conducted and it has mainly been identified based on its morphology. In this study, the Korean specimens considered to be A. auricula-judae based on their morphology were identified as three differnt species (A. heimuer, A. minutissima, and A. villosula) phylogenetically. Furthermore, the absence of A. auricula-judae in Korea was confirmed in our ITS analysis (Fig. 1). The geographical distribution of A. auricula-judae has previously been confined to Europe [4-6]. A phylogenetic tree confirmed the phylogenetic relationships of Korean Auricularia species for the first time.
The cultivated strains of A. auricula-judae from Korea were sequenced and analyzed phylogenetically (data not shown). As a result, the strains cultivated as products in the markets were identified as A. heimuer, which has a wide geographic distribution in East Asia. Auricularia auricula-judae. s. str. is not found in China, Japan, or the Russian Far East based on geological distribution [4], and this species is confined to European countries [4,7].
Auricularia hispida was introduced as a new species in Japan (Index Fungorum; https://www.indexfungorum.org/). This species was excluded in a recent Japanese taxonomical study [6], and there is no sequence data available for it in GenBank. In Korea, we could not find this species during the field surveys. Auricularia mesenterica is a type species of Auricularia and is considered a species complex [4]. A. mesenterica complex was split into three species (A. mesenterica s.str., A. brasiliana, and A. orientalis) based on phylogenetic analysis. Auricularia mesenterica s.str. is a European species, A. orientalis was found in China, and A. brasiliana was discovered in Brazil. We could not identify these species in Korea. Therefore, more samples are required to identify this species. Auricularia polytricha (Mont.) Sacc. was recorded in Korea, and the species is synonymized with A. nigricans (Sw.) Birkebak, Looney & Sánchez-García. According to a recent phylogenetic analysis [6], this species was recorded in the USA and Costa Rica, however it was not identified in this investigation. Therefore, additional collections are required to confirm its presence in Korea.

