서론
기수역(brackish water)은 담수와 해수가 혼합되어 형성되는 지역으로 일반적으로 염분(NaCl)의 농도가 0.05%이하인 물은 담수(freshwater), 3%이상은 해수(seawater)라 하는데 중간을 기수(brackish)라 정의한다[1]. 미생물이 실제 이용할 수 있는 물의 양은 수분활성도(water acivity, aw)로 나타낼 수 있는데, 증류수의 수분활성도는 1이 되며, 염분농도는 수분활성도를 감소시킨다. 해양에서 분리되는 미생물은 생장에 염분을 필요로 하는데 바닷물의 수분활성도에서 최적 생장하는 미생물을 호염성 미생물(halophile)이라 한다. 이 중 15-30%의 매우 높은 농도에서 생장 가능한 미생물은 극호염성미생물(extreme halophile)이라 한다. 균류 중에서는 대표적으로 Saccharomyces cerevisiae, Debaryomyces hansenii 등이 호염성 효모로 보고되어 있다[2].
본 연구에서는 국내 기수 환경의 물과 토양에서 균류를 분리 및 동정하여 지역에 따른 균류의 분포와 다양성지수를 확인하였다. 그리고 이들 중 국내 미기록 균류 2종을 선발하여 종 특성을 분석하였다.
재료 및 방법
시료 채집 및 균류 분리
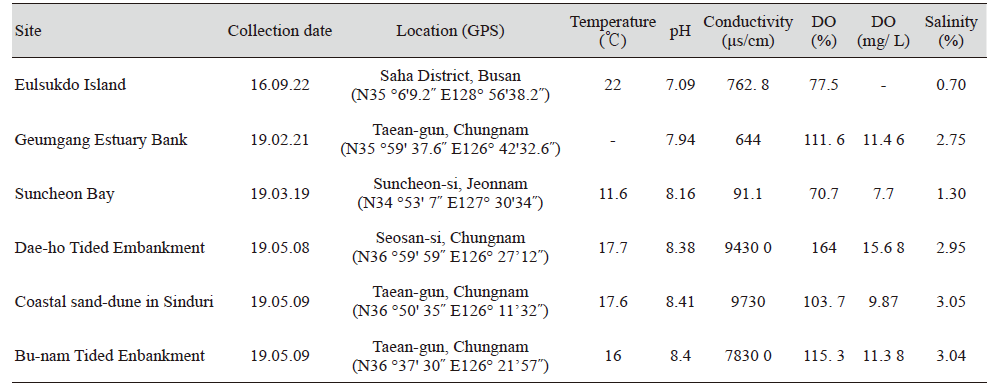
부산 사하구 을숙도, 충남 태안군 금강하구둑, 전남 순천시 순천만습지, 충남 서산시 대호방조제, 태안군 신두리사구 및 부남방조제 기수지역에서 물과 퇴적물을 채집하여 균류를 분리하였다(Table 1). 조사지점의 물시료를 50 mL 채취하여 현장에서 핸드펌프와 nitrocellulose membrane filter (pore size 0.45 µm MCE membrane, MF-MilliporeTM, Darmstadt, Germany)를 이용하여 여과하였다. 필터의 상면을 water agar (WA; 20g/L, agar) 각각 부착하여 15℃에서 1일간 배양 후, membrane filter를 제거하고 실체현미경(Stemi 305, Carl Zeiss, CN, Germany)을 이용하여 발아한 포자를 배지에서 분리하고 V8배지(V8A; 8% V8 juice [v/v] and 1.5 agar [w/v] adjusted to pH6.0 using 10 N NaOH)에 배양하였다. 채집된 토양시료는 101, 102, 103으로 희석하여 50 ppm 항생제(streptomycin)가 첨가된 potato dextrose agar (PDA; Difco, Detroit, MI, USA)에 도말한 후 3-4일 동안 15℃에서 배양하였다. 배양된 배지에서 단포자 분리를 통해 V8 배지에 순수분리하였다. 순수분리한 균류는 PDA에 계대배양하였다.
균류의 동정 및 계통수 작성
분리된 균주들의 ITS (internal transcribed spacer) 부위의 염기서열을 분석하기 위해 ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) 및 ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) primer를, RPB2 유전자분석을 위해서는 5F2 (5’-GGGGWGAYCAGAAGAAGGC-3’) 및 7cR (5’-CCCATRGCTTGYTTRCCCAT-3’) primer를 이용하여 분석한 후 NCBI의 BLAST를 사용하여 데이터베이스상의 기등록 균류들과의 상동성을 비교하였다[3,4]. 국내에 기록되지 않은 균류의 계통수 작성은 MEGA7을 이용하였다[5].
또한, 형태적 특징을 관찰하기 위해 광학현미경(H550S, Nikon, Yokohama, Japan)을 이용하여 화상자료를 확보하였다.
균류의 생장 특성 분석
국내 미기록 균류의 배지별 생장길이를 측정하기 위해 PDA, malt extract agar (MEA; 2% malt extract [w/v], 2% agar [w/v]), oatmeal agar (OA; Difco, Detroit, MI, USA), yeast extract peptone dextrose agar (YPDA; Difco, Detroit, MI, USA), 그리고 1x sea water PDA (1xSPDA, 2.4% NaCl [w/v], 0.7% MgSO4·7H2O [w/v], 0.53% MgCl2·6H2O [w/v], 0.07% KCl [w/v], 0.013% CaCl2·2H2O [w/v], PDA), 2x sea water PDA (2xSPDA, 4.8% NaCl [w/v], 1.4% MgSO4·7H2O [w/v], 10.6% MgCl2·6H2O [w/v], 0.14% KCl [w/v], 0.026% CaCl2·2H2O [w/v], PDA), 4x sea water PDA (4xSPDA, 9.6% NaCl [w/v], 2.8% MgSO4·7H2O [w/v], 2.12% MgCl2·6H2O [w/v], 0.28% KCl [w/v], 0.052% CaCl2·2H2O [w/v], PDA) 고체배지에 5, 15, 20, 25, 30℃에서 20일동안 배양하였다.
다양성 지수
Shannon diversity index (H)는 community 안에서 종 다양성을 보는 지수로서, abundance와 evenness를 동시에 수치화한다. 수치가 높아질수록 종이 다양하게 분포하고 있음을 나타낸다[6]. Shannon’s equitability (EH)는 H를 Hmax (lnS) 값으로 나눈것으로서, 0과 1사이 값이 검출되며, 1은 완벽한 evenness를 나타낸다[6]. Shannon’s evenness index (SEI)는 H를 최대값으로 나눈 것으로서 군집에서의 상대적인 abundance를 나타낸다[6](Table 2).
Simpson’s diversity index (D)는 서식지 내에서의 종 다양성을 정량화하는데 주로 사용한다. 존재하는 종의 수와 각 종의 abundance를 나타낸다. D의 값은 0과 1사이이며, 0은 무한한 다양성을 나타내고, 1은 다양성이 없음을 보여준다[7](Table 2).

결과
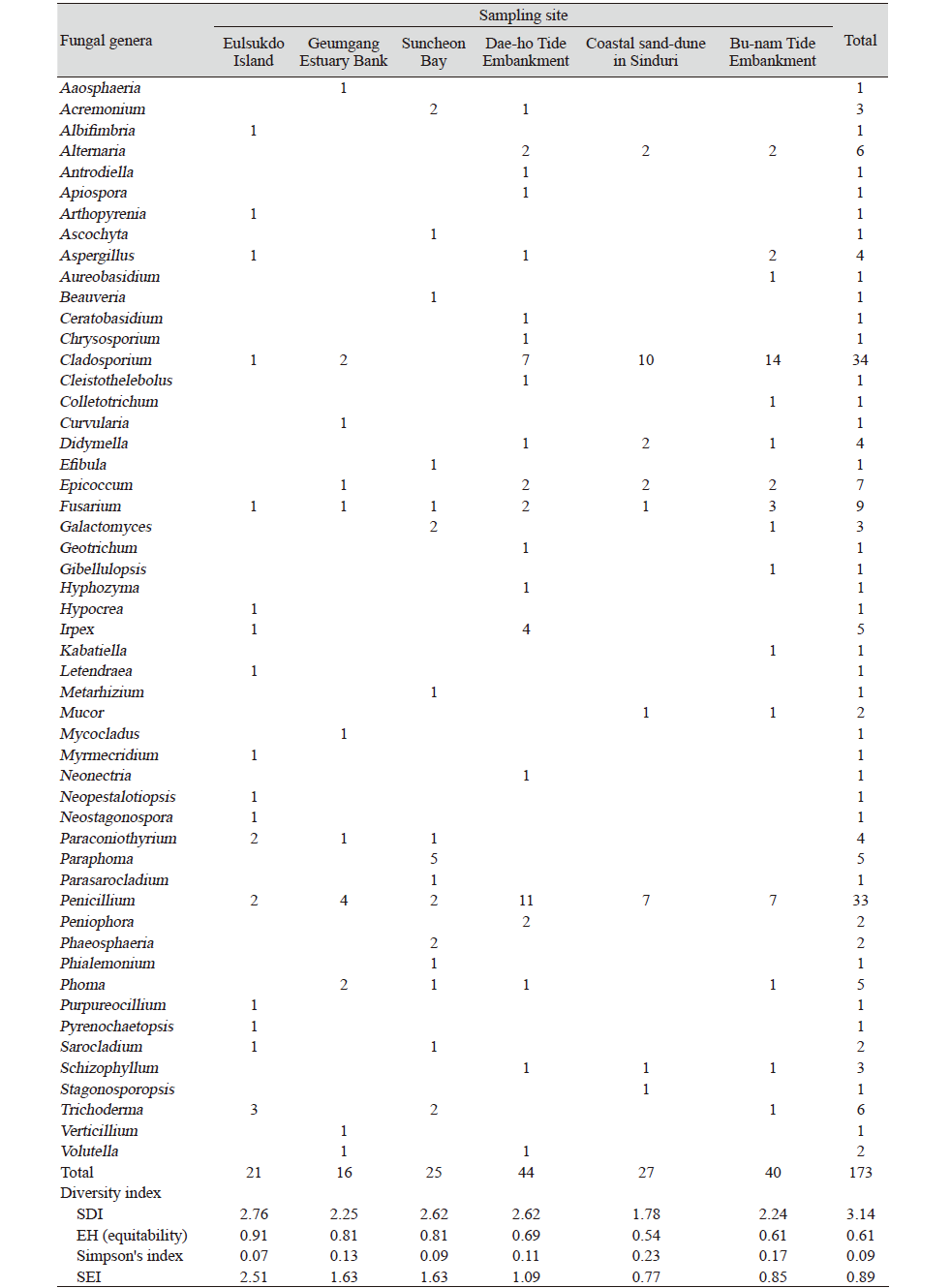
국내 기수역 중 6개의 지역으로부터 을숙도에서 21 균주, 금강하구둑에서 16균주, 순천만습지에서 25균주, 대호방조제에서 44균주, 신두리사구에서 27균주, 부남방조제에서 40균주로 총 173균주를 분리, 동정하였다. NCBI의 GenBank로부터 모든 균주에 대하여 등록번호를 부여받았다(Table 3). 각 지역에서 분리된 균류의 다양성은 Table 4에 나타난 바와 같다.
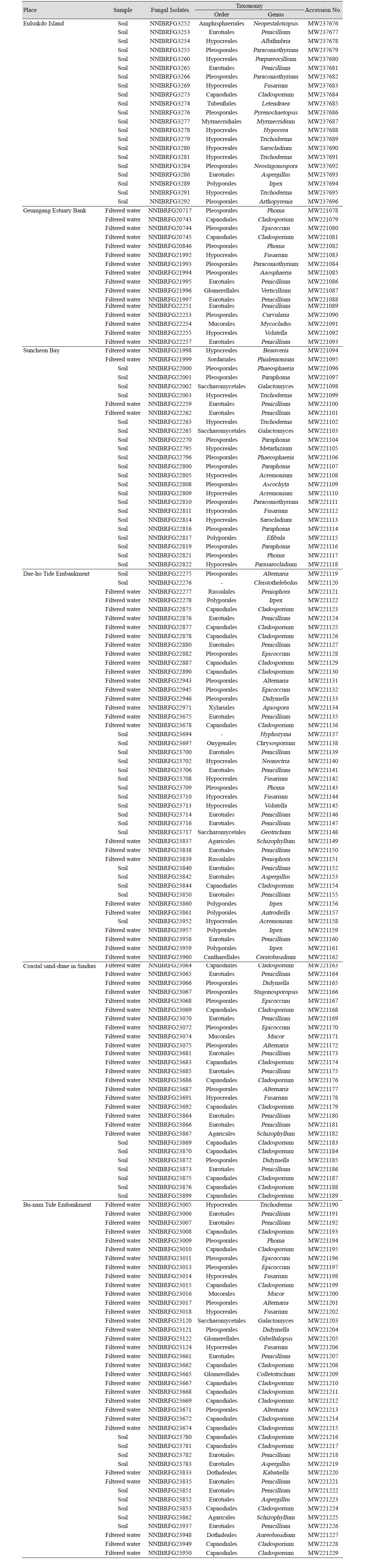
Table 4. Diversity index and distribution of fungi isolated brackish water.
|
|
* SDI: Shannon's divsersity index, SEI: Shannon's evenness index |
을숙도에서 분리된 21개 균주 중 균류는 Ascomycota문이 대부분이었고 1개 균주가 Basidiomycota문 Polyporales목의 Irpex속에 속했다. Ascomycota문에 속하는 균주는 Capnodiales목에 속하는 Cladosporium속과 Pleosprales목에 속하는 Arthopyrenia속, Paraconiothyrium속, Neostagonospora속, Pyrenochaetopsis속, Tubeufiales목의 Letendraea속, Eurotiales목의 Aspergillus속, Penicillium속, Amphisphaeriaels목의 Neopestalotiopsis속, 그리고 Hypocreales목의 Hypocrea속, Trichoderma속, Fusarium속, Purpureocillium속, Sarocladium속, Albifimbria속, Myrmecridium속으로 확인되었으며 Hypocreales목이 38% (8균주)로 가장 많은 비중을 차지하였다(Table 4).
금강하구둑에서 분리된 균류 또한 Ascomycota문이 대부분이었고 1개 균주가 Mucoromycota문 Mycocladus속에 속했다. Asocomycota문에 속하는 균주는 Capnodiales목 Cladosporium속과 Pleosprales목의 Aaosphaeria속, Phoma속, Epicoccum속, Paraconiothyrium속, Curvularia속, Eurotiales목의 Penicillium속, Glomerellales목의 Verticillium속, Hypocreales목의 Fusarium속, Volutella속으로 확인되었으며 Eurotiales목의 Penicillium속이 25%(4균주)로 가장 많은 비중을 차지하였다(Table 4).
순천만습지에서 분리된 균류는 Ascomycota문이 대부분이었고 1개 균주가 Basidiomycota문Polyporales목의 Efibula속에 속했다. Asocomycota문에 속하는 균주는 Pleosporales목의 Ascochyta속, Phoma속, Paraconiothyrium속, Paraphoma속, Phaeosphaeria속, Eurotiales목의 Penicillium속, Saccharomycetales목의 Galactomyces속, Hypocreales목의 Acremonium속, Metarhizium속, Beauveria속, Trichoderma속, Fusarium속, Parasarocladium속, Sarocladium속, Phialemonium속으로 확인되었으며 Pleosporales목이 40% (10균주)로 가장 많은 비중을 차지하였다(Table 4).
대호방조제에서 분리된 균류는 Ascomycota문 및 Basidiomycota문에 속했으며 Ascomycota문에 속하는 균주는 Cleistothelebolus속, Hyphozyma속과 Capnodiales목의 Cladosporium속, Pleosporales목의 Phoma속, Didymella속, Epicoccum속, Alternaria속, Eurotiales목의 Aspergillus속, Penicillium속, Onygenales문의 Chrysosporium속, Saccharomycetales문의 Geotrichum속, Hypocreales문의 Acremonium속, Fusarium속, Neonectria속, Volutella속, Xylariales문의 Apiospora속이 확인되었고 Basidiomycota문에 속하는 균주는 Cantharellales목의 Ceratobasidium속, Agaricales목의 Schizophyllum속, Polyporales목의 Antrodiella속, Irpex속, Russulales목의 Peniophora속이 확인되었다. 그 중 Eurotiales목의 Penicillium속이 25%(11균주)로 가장 많은 비중을 차지하였다(Table 4).
신두리사구에서 분리된 균류는 Ascomycota문이 대부분이었고 Basidiomycota문 Agaricales목의 Schizophyllum속, Mucoromycota문 Mucorales목의 Mucor속에 각각 1개 균주씩 속했다. Ascomycota문에 속하는 균주는 Capnodiales목의 Cladosporium속, Pleosporales목의 Didymella속, Epicoccum속, Stagonosporopsis속, Alternaria속, Eurotiales목의 Penicillium속, Hypocreales목의 Fusarium속이 확인되었고 Capnodiales목의 Cladosporium속이 37%(10균주)로 가장 많은 비중을 차지하였다(Table 4).
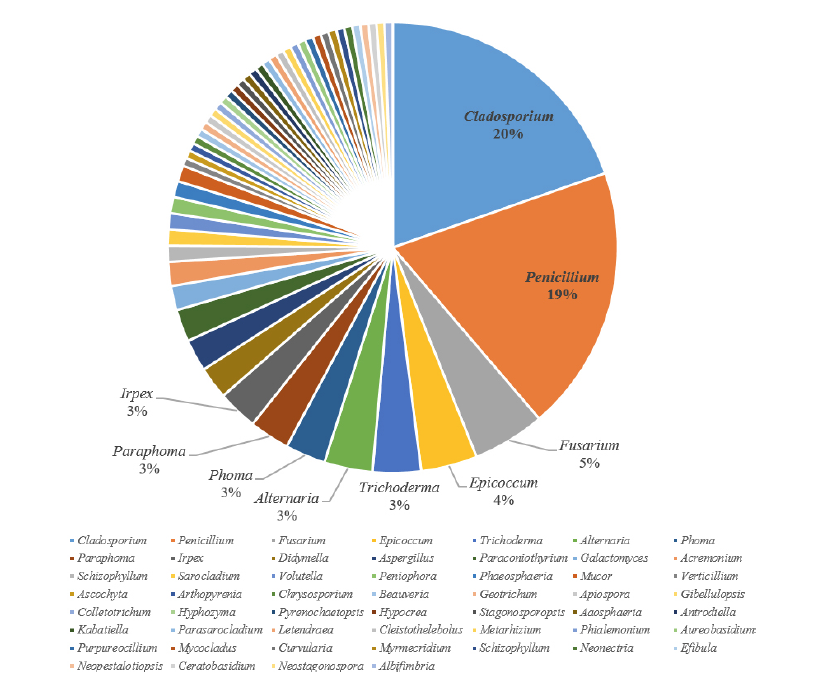
부남방조제에서 분리된 균류 또한 Ascomycota문이 대부분이었고 Basidiomycota문 Agaricales목의 Schizophyllum속, Mucoromycota문 Mucorales목의 Mucor속에 각각 1개 균주씩 속했다. Ascomycota문에 속하는 Capnodiales목의 Cladosporium속, Dothideales목의 Aureobasidium속, Kabatiella속, Pleosporales목의 Phoma속, Didymella속, Epicoccum속, Alternaria속, Eurotiales목의 Aspergillus속, Penicillium속, Saccharomycetales목의 Galactomyces속, Glomerellales목의 Collectotrichum속, Gibellulopsis속, Hypocreales목의 Trichoderma속, Fusarium속이 확인되었고 Capnodiales목의 Cladosporium속이 35% (14균주)로 가장 많은 비중을 차지하였다. 6개의 기수역에서 Asocomycota문이 90.8% (157균주)로 대부분을 차지하였고, 그 중에서 Capnodiales목의 Cladosporium속이 19.7% (34균주)로 가장 많았고, Eurotiales목의 Penicillium속이 19%(33균주)로 그 다음 비중을 차지하였다(Table 4).
Taxonomy
Sarocladium kiliense (Grütz) Summerbell, Studies in Mycology 68: 158 (2011) [MB#519592]
(Fig. 1 and Fig. 2)
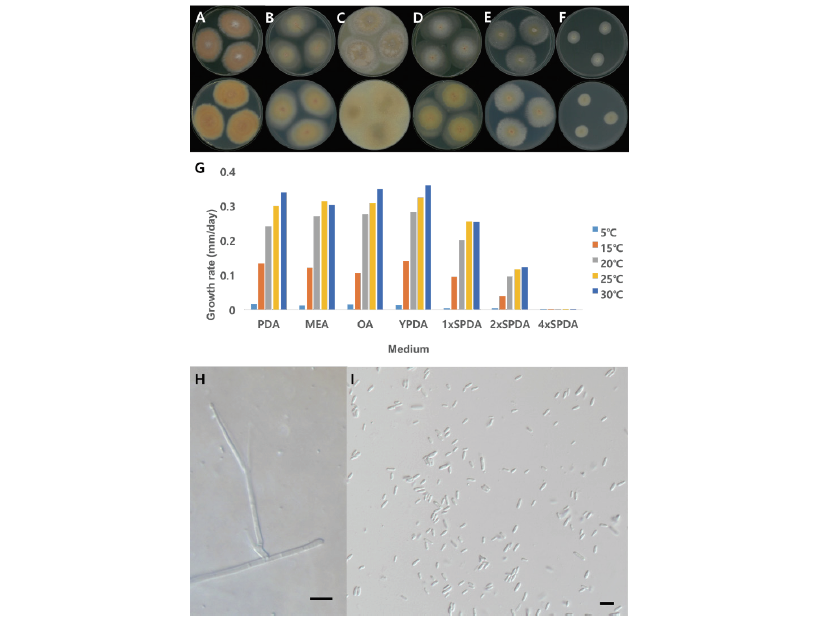
Fig. 1.Cultural and morphological characteristics of NNIBRFG3280. Macro-morphology of front and back. A: Potato dextrose agar (PDA), B: Malt extract agar (MEA), C:Oatmeal agar (OA), D: Yeast extract peptone dextrose agar (YPDA), E: 1x sea water PDA (1xSPDA), F: 2x sea water PDA (2xSPDA), G: Growth rate on media. Micro-morphology of conidiophore (H) and conidia (I). Size bar is 10 μm.
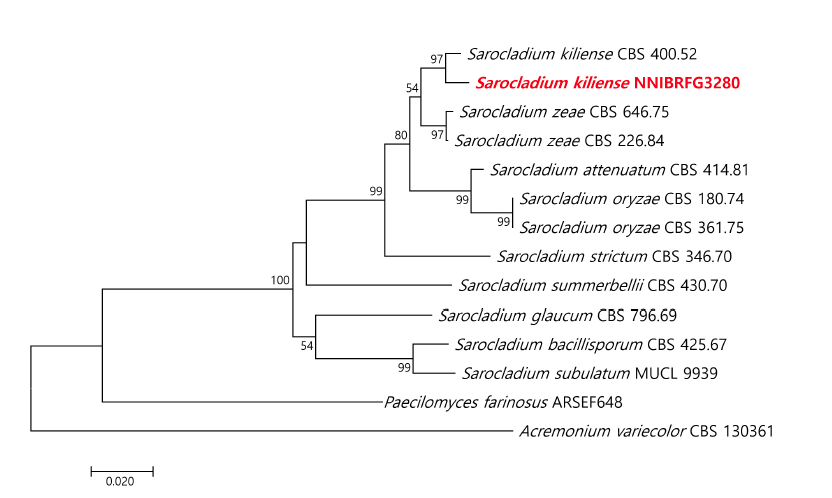
Fig. 2.Phylogenetic tree of Sarocladium kiliense NNIBRFG3280 and closely related species based on a neighbor-joining analysis of the internal transcribed spacer (ITS) sequences. The sequence of Paecilomeces farinosus and Acremonium variecolor were used as an outgroup. Numbers at the nodes indicate the bootstrap values (>50%) from 1,000 replications. The bar indicates the number of substitutions per position. The new isolate from the present study is shown in bold and red.
Characterization: NNIBRFG3280균주는 PDA배지에 배양했을 때 30℃에서 0.34 mm/day 정도로 느리게 생장하였다. PDA배지에서 균사체는 가운데 부분이 흰색을 띄다가 중간부분은 연한 분홍색이고 가장자리 부분은 흰색을 띄었다(Fig. 1A). MEA, OA, YPDA 배지에서는 콜로니의 가운데 부분은 분홍색을 띄고 중간은 연한 노란색을 가장자리 부분은 흰색을 띄었다(Fig. 1B-D). 대체로 30℃에서 잘 생장하였으며, YPDA배지에서 가장 높은 생장율을 보였다. 염분농도가 높은 4xSPDA에서는 생장하지 않았다(Fig. 1G).
균사는 투명하고 매끄러운 표면을 가지며 너비는 1-3 μm이다. Phialides는 균사에서 개별적으로 뻗어 있었으며 길이가 20-41 μm, 너비가 1-2.5 μm이었다. 분생포자(conidia, n=20)는 길쭉한 원통형이거나 타원형의 모양이며 매끄러운 표면을 가지고 있었고 길이가 4-7 μm, 너비는2-3 μm이었다(Fig. 1H and Fig. 1I).
Habitat: 기수역의 퇴적물
Specimen examined: 부산시 사하구 을숙도, 2016.09.22., NNIBRFG3280, 국립낙동강생물자원관
Note: 1925년 Grütz에 의해서 Acremonium kiliense로 처음 보고되었으며[8] 2011년 Summerbell에 의해 Sarocladium kiliense로 다시 명명되었다[9]. Summerbell 등[9]이 보고한 바에 의하면 Sarocladium은 Acremonium과 달리 길쭉한 phialides를 가지며 adelophialides를 많이 생산하고 길쭉한 분생자를 가지는 형태적 특징이 있다. Sarocladium 속엔 10종이 속해 있으며 그 중 S.kiliense는 배지별 다른 콜로니 색을 띄는 특징을 가지는데 NNIBRFG3280 균주 또한 같은 특징을 보였다. NNIBRFG3280균주의 ITS영역을 blast한 결과, S. kiliense (MT000479)와 99.79%의 높은 유사도를 보였으며 계통수에서도 S. kiliense와 같은 clade에 위치하여 최종적으로 S. kiliense로 동정하였다. NNIBRFG3280균주는 염분 0.7%인 기수역의 퇴적물에서 분리하였으며 30℃에서 염분 6%인 2xSPDA에서 생장하는 것으로 보아 염분내성이 있는 균류로 확인되었다(Fig. 1F and Fig. 1G).
Fusicolla merismoides (Corda) Gräfenhan, Seifert & Schroers, Studies in Mycology 68: 101 (2011) [MB#519438](Fig. 3 and Fig.4)
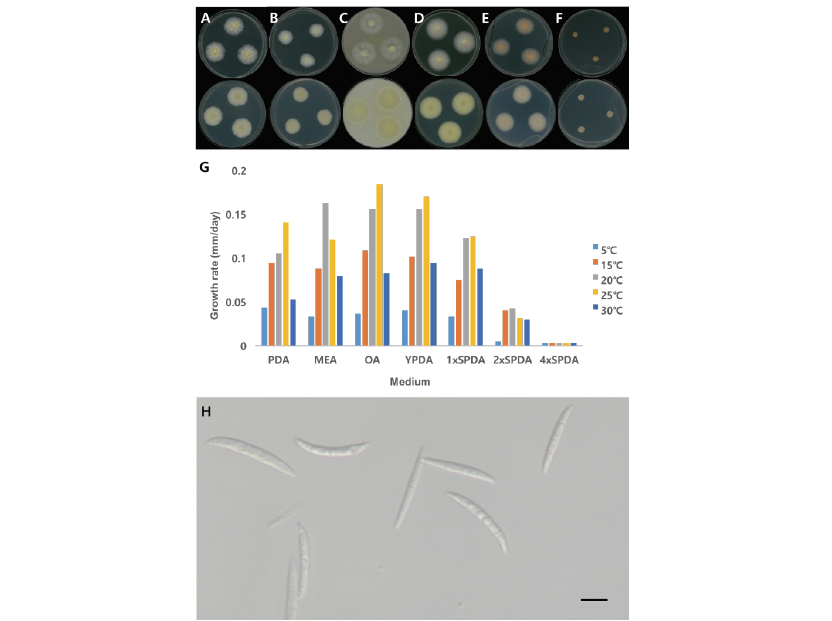
Fig. 3.Cultural and morphological characteristics of NNIBRFG23708. Macro-morphology of front and back. A: Potato dextrose agar (PDA), B: Malt extract agar (MEA), C: Oatmeal agar (OA), D: Yeast extract peptone dextrose agar (YPDA), E: 1x sea water PDA (1xSPDA), F: 2x sea water PDA (2xSPDA), G: Growth rate on media. H: Micro-morphology of conidia. Size bar is 10 μm.
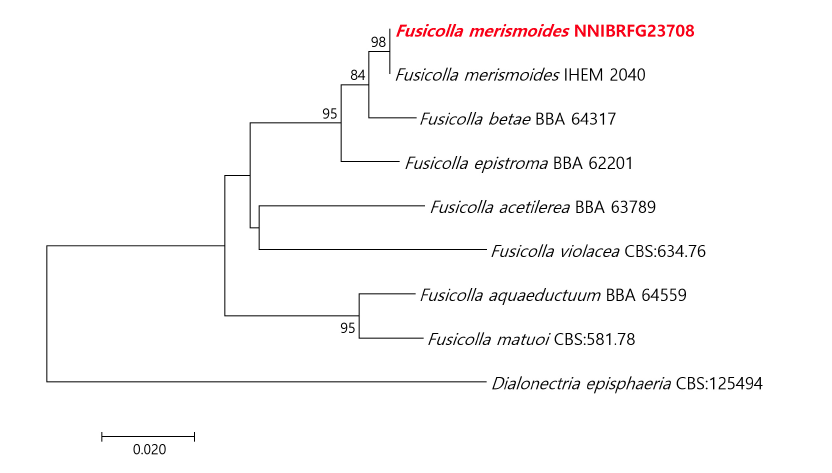
Fig. 4.Phylogenetic tree of Fusicolla merismoides NNIBRFG23708 and closely related species based on a neighbor-joining analysis of RPBS2 gene sequences. The sequence of Dialonectria episphaeria was used as an outgroup. Numbers at the nodes indicate the bootstrap values (>50%) from 1,000 replications. The bar indicates the number of substitutions per position. The new isolate from the present study is shown in bold and red.
Characterization: NNIBRFG23708균주는 PDA배지에 배양했을 때 25℃에서 0.14 mm/day 정도로 느리게 생장하였다. PDA배지에서 균사체는 가운데 부분이 옅은 노란색을 띄다가 가장자리 부분은 흰색을 띄었다(Fig. 3A). MEA, OA, YPDA 배지에서도 PDA배지에서와 비슷한 양상을 보였으며(Fig. 3B-D), 염분이 포함된 배지에서는 옅은 분홍색을 띄었다(Fig. 3E and Fig. 3F). 대체로 25℃에서 잘 생장하였으며, 염분농도가 높은 4xSPDA에서는 생장하지 않았다(Fig. 3G).
Microconidia는 관찰되지 않았고 macroconidia의 길이는 145-228 μm이고 너비는 15-26 μm이며1-3개의 격막을 가지고 있고 길쭉한 모양으로 끝이 뾰족하고 약간 휘어져있다(Fig. 3H).
Habitat: 기수역의 퇴적물
Specimen examined: 충청남도 서산시 대호방조제, 2019.05.08., NNIBRF23708, 국립낙동강생물자원관
Note: Fusicolla merismoides는 보통 토양과 오염된 물에서 분리되었으며 1838년 Corda에 의해서 Fusarium merismoides로 처음 보고되었다[10]. NNIBRFG23708균주의 ITS염기서열을 NCBI에서 blast한 결과, Fusarium merismoides (MT072065)와 100%의 유사도를 보였다. 이 균주를 최종 동정하기 위해서 ITS영역으로는 확인이 불가능하여 RPB2 유전자를 분석하여 계통수를 작성한 결과, Fusicolla merismoides와 같은 clade에 묶이는 것을 확인할 수 있었다. NNIBRFG23708균주는 염분 2.95%인 기수역의 퇴적물에서 분리되었으며 25℃에서 염분 6%인 2xSPDA에서 생장하는 것으로 보아 염분내성이 있는 균류로 확인되었다(Fig. 3F and Fig. 3G).
고찰
염분이 있는 인도의 맹그로브 숲과 solar saltern의 물 및 토양으로부터 Aspergillus 속 및 Penicillium속이 주로 검출되었고[11], 오클라호마의 Great Salt Plains호수의 토양에서는 Aspergillus 속 및 Penicillium 속 등이 분리되었다[12]. 우리나라의 고창과 무안갯벌에서는 Penicillium 속이 가장 많았고, 그 다음으로 Fusarium 속 및 Aspergillus 속이 많았다[13,14]. 본 연구에서는 목 수준에서는 Pleosporales 목, Eurotiales 목, Capnodiales 목이 가장 많이 검출되었고, 속 수준에서는 Cladosporium 속 및 Penicillium 속이 우점하는 것으로 나타나 다른 연구결과와 비슷한 양상을 보였다(Fig. 5). 각 기수지역으로부터 다양성지수를 도출한 결과, Shannon’s diversity index와 Simpson’s index 모두 염분이 낮게 측정된 을숙도 지역에서 가장 다양성이 높게 나왔으며, 염분이 제일 높았던 신두리사구 지역에서는 낮게 나왔다(Table 3). 이는 염분에 내성이 있는 균류의 다양성이 높지 않음을 보여준다.
한편 Nayak 등[11]의 연구에서는 곰팡이의 순수분리시에 염분이 함유된 배지를 사용하여 절대호염성(obligate halophile) 곰팡이를 분리할 수 있었으나, 본 연구에서는 일반 PDA에 배양하여 통성호염성(facultative halophile) 곰팡이만을 분리할 수 있었다. 국내 미기록으로 확인된 두 균주 역시 넓은 범위의 염분내성을 가지고 있지만 염분 배지에서 최적 생장을 하지 않는 것으로 보아 통성호염성 곰팡이임을 알 수 있다(Fig. 1 and Fig. 3).
적 요
본 연구에서는 국내 기수 환경의 물과 토양에서 균류의 다양성을 확인하기 위해 을숙도, 금강하구둑, 순천만습지, 대호방조제, 신두리사구 및 부남방조제 총 6지점의 현장조사 및 시료를 채집하였다. 현장에서 여과한 담수시료와 담수퇴적물로부터 곰팡이를 순수분리하여 PDA배지에 접종한 후 25℃에서 배양하였다. 기수역 시료에서 순수분리된 균주는 총 173균주로 ITS 부분을 분석하여 속 수준으로 동정하였다. 분리 동정된 균주를 다양성 지수를 이용하여 분석하였다. 분리된 균류는 Pleosporales, Eurotiales, Capnodiales, Hypocreales, Polyporales, Saccharomycetales, Agaricales, Glomerellales, Mucorales, Dothideales, Russulales, Xylariales, Sordariales, Myrmecridiales, Tubeufiales, Onygenales, Cantharellales, Amphisphaeriales 목에 속하며 Cladosporium속이 20%로 가장 우점하며, 그 다음은 Penicillium (19%), Fusarium (5%)의 순서로 많이 분포하였다. Sarocladium kiliense NNIBRFG3280와 Fusicolla merismoides NNIBRFG23708를 국내 기수환경에서 발굴하였으며, 이를 국내 미기록종으로 보고하고자 한다.

