Introduction
The Inocybaceae is a family of ectomycorrhizal fungi that is estimated to contain 1,050 species [1]. This family is associated with 23 families of vascular plants, including Betulaceae, Dipterocarpaceae, Fagaceae, Pinaceae, and Salicaceae, in mycorrhizal symbiosis [1,2]. In addition, many species are known to be poisonous mushrooms and produce secondary metabolites, such as muscarine, psilocybin, and other compounds. The genus Inocybe has been divided by different authors into varying numbers of subgenera, sections, and subsections based on morphological characteristics [3-5]. Recently, seven genera that establish a global overview of multigene molecular phylogenetic analyses in the Inocybaceae have been presented [6]: Auritella Matheny & Bougher, Inocybe (Fr.) Fr., Inosperma (Kühner) Matheny & Esteve-Rav., Mallocybe (Kuyper) Matheny, Vizzini & Esteve-Rav., Nothocybe Matheny & K.P.D. Latha, Pseudosperma Matheny & Esteve-Rav., and Tubariomyces Esteve-Rav. & Matheny. To date, 32 Inocybaceae species have been listed in Korea (Index of Korean Mushroom, ) [7], but Inocybaceae diversity remains relatively unexplored in recent decades. A few studies have reported taxonomic identification [8-10], all of which were based on morphology. Since 2008, approximately 350 Inocybaceae specimens have been collected and preserved in the Korea National Arboretum (KH, Pocheon, Korea). Here, based on morphological characterization and phylogenetic analysis using internal transcribed spacer (ITS) DNA barcodes, seven species of Inocybaceae were newly confirmed in Korea.
Materials and Methods
Morphological observation
A total of 350 samples were collected during the 2008-2020 survey. Of these, 13 samples were examined for identification based on macroscopic and microscopic characteristics in the present study (Table 1). Dried materials were mounted in distilled water and 5% KOH using an Olympus BX53 microscope (Olympus Corporation, Tokyo, Japan) and Jenoptik ProgRes C14 Plus Camera (Jenoptik Corporation, Jena, Germany). Microscopic parameters were measured using ProgRes Capture Pro v.2.8.8. software (Jenoptik Corporation, Jena, Germany). The systematics of the taxa used in this study were in accordance with the Index Fungorum (). Dried specimens were deposited in the herbarium of the Korea National Arboretum (KH).
DNA extraction and phylogenetic analysis
For phylogenetic analysis, genomic DNA from the specimens was extracted using a DNeasy Plant Mini DNA Extraction Kit (Qiagen, Inc., Valencia, CA, USA). The ITS regions of rDNA were amplified with primers ITS5 and ITS4 [11]. The polymerase chain reaction amplicons were purified using a QIAquick Purification Kit (Qiagen, Inc., Valencia, CA, USA) and directly sequenced using an ABI Prism 377 Automatic DNA Sequencer (Applied Biosystems, Foster City, CA, USA) with a BigDyeTM Cycle Sequencing Kit (version 3.1, Applied Biosystems, Foster City, CA, USA). GenBank accession numbers are listed in Table 1. The sequences were incorporated with those of closely related species retrieved from a public database (National Center for Biotechnology Information; https://www.ncbi.nlm.nih.gov/). The dataset was aligned using MAFFT v.7 [12]. All other parameters were set to the default values. A phylogenetic tree was constructed using RAxML in the CIPRES Science Gateway (). Crepidotus malachioides (NR 132047) was used as an outgroup species. The relative robustness of the individual branches was estimated by bootstrapping with 1,000 replicates.
Results and Discussion
Phylogenetic results
Among the 350 specimens collected, 13 of them were found to be unrecorded species in Korea. Six species of Inocybaceae (Inocybe fuscidula, Inocybe grammatoides, Inocybe latibulosa, Inocybe stellata, Inosperma shawrense, and Mallocybe malenconii) formed well-supported clades with 100% bootstrap values in the ITS-based phylogeny (Fig. 1). Inocybe caroticolor formed a monophyletic clade with 99% support. The sequence of I. stellata from Korea grouped into I. stellata but was divided into three clades. One clade contained the type species (NR_153122) for I. stellata from Thailand. Horak et al. [13] showed that this species can be divided into two clades in nLSU phylogeny. In addition, rpb2 sequences also supported the distinction between the two clades. Therefore, the Korean specimen (MW520962) was temporarily identified as I. stellata. Additional samples and multigene analyses will help in future identification.
Taxonomy
Inocybe caroticolor T. Bau and Y.G. Fan, Mycotaxon 123: 170, 2013 (Figs. 1 and 2)
Korean name: Juhwang-Ttambeoseot (주황땀버섯); derived from the color of pileus; orange color.
Description: Pileus 15-30 mm in diameter, conical when young, then convex, with an obtuse umbo, radially fibrillose, rimose. Lamellae light yellow to yellowish orange, regularly arranged, adnexed, and slightly crowded. Stipe 20-32×2-4 mm, central, cylindrical, pruinose entire length, densely at apex, longitudinally striate, and reddish orange at apex, basal tomentum white. Basidiospores 6.5-9.5×4.7-6 µm, pale brown, weakly to strongly nodulose. Basidia 24-37×5.5-7 µm, clavate, 4-spored, with yellowish pigments. Pleurocystidia 48-60×11-14 µm, fusiform, obtuse base. Cheilocystidia 47-60×12-14.8 µm, fusiform, some cylindrical. Caulocystidia 49-75×9-13 µm, variably shaped, abundant, thin-walled. Pileipellis 0.2-6 µm, a cutis of cylindrical hyphae, smooth, pigmented, thin-walled. Stipitipellis 2-4.5 µm, a cutis composed of regularly arranged hyphae, filamentous, smooth, thin- to moderately thick-walled. Clamp connections present.
Note: In 2013, this species was proposed as a new species in China [14]. It is characterized by its carrot-colored basidiomata, entirely pruinose stipe, aromatic odor, and nodulose basidiospores. Morphological examination and phylogenetic analysis confirmed the presence of this species in Korea.
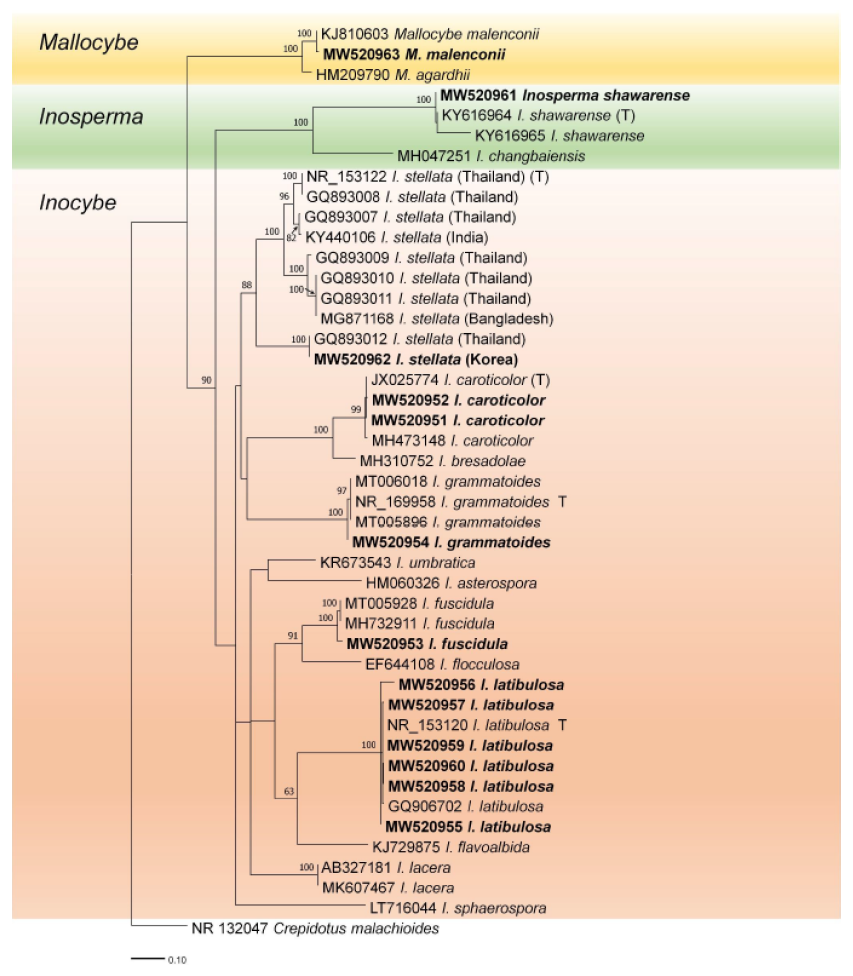
Fig. 1. Randomized Axelerated Maximum Likelihood (RaxML) tree of the Inocybaceae family using internal transcribed spacer (ITS) regions of rDNA sequences. Crepidotus malachioides is an outgroup. Bootstrap values higher than 60% are shown in the branches (1,000 replicates). The scale bar equals the number of nucleotide substitutions per site. The type materials are indicated as T in phylogeny. Our specimens are named in bold.

Inocybe fuscidula Velen., České Houby 2: 378, 1920 (Figs. 1 and 3)
Korean name: Damgalsaekkkokji-Ttambeoseot (담갈색꼭지땀버섯); derived from the characterization of pale brown and distinctly umbonate pileus.
Description: Pileus 4.5-5.5×0.8-1.2 cm, when young conical convex, flatly convex to convex, also applanate, umbonate, margin when young inflexed and markedly appendiculate with a whitish fugacious cortina, when old expanding, rimose. Gills with intermediates, 0.4-0.6 cm broad, from flat to strongly arched, emarginated, generally adnate for a quarter, occasionally semi-adnate, almost free, when young whitish, gray-beige, when old dark beige, when old dark beige to light brownish, edges even, flocculose. Stipe 7-8×0.3-0.8 cm, cylindrical, parallel, often compressed, bent, with slightly swollen to slightly bulbous base covered with whitish mycelial strands, the upper third distinctly pruinose, finely fibrillose towards the base. Basidia 25-30×8-10 µm, generally 4-spored. Basidiospores 8-11×4.5-6 µm. Cheilocystidia 35-70×16-23 µm. Pleurocystidia 45-75×13-22 µm, walls 1.5-2 µm thick. Caulocystidia from the upper stipe 38-65×13-22 µm. Cap cuticle hyphae 8-12 µm wide.
Note: Inocybe fuscidula is a well-known species worldwide. In the present study, we confirmed this species based on morphology and ITS sequence.

Inocybe grammatoides Esteve-Rav., Pancorbo & E. Rubio, in Crous et al., Persoonia 42: 419, 2019 (Figs. 1 and 4)
Korean name: Huingaru-Ttambeoseot (흰가루땀버섯); derived from the pruinose stipe; covered with white powdery granules.
Description: Pileus 15-55 mm, at first conical-campanulate, then convex to plano-convex, broadly umbonate to subumbonate, margin straight, regular to hardly wavy with age, color pinkish gray when young or moistened, to light gray or very pale brown when drying, uniform, surface radially fibrillose, smooth. Lamellae moderately crowded, adnexed to emarginate, ventricose, initially whitish, becoming pale gray to light brown, edge paler to concolorous. Stipe 30-65×5-10 mm, straight to curved towards base, cylindrical, clavate to subbulbous, whitish becoming beige to ochraceous towards the lower half with age, surface densely pruinose at the upper half, becoming sparsely pruinose toward the base. Context fibrose, whitish, pinkish in the upper part of the stipe. Basidiospores 7.0-9.5×4.5-7.0 µm, polygonal-subrectangular yellowish. Basidia 27-37×7.5-10 µm, 4-spored, clavate, sterigmata 3.5-6 µm long. Pleurocystidia abundant, 48-80×10.5-23 µm, fusiform, hyaline, pale to moderately yellowish. Cheilocystidia similar in size and shape to pleurocystidia. Stipitipellis a cutis bearing numerous caulocystidia, more scattered towards the base, mixed with clavate to broadly clavate hyaline paracystidia. Pileipellis parallel cylindrical cells, showing minute pale intracellular pigment. Clamp connections present.
Note: This species was proposed as a new species in 2019 [15]. This species behaves as a mesophilic species, usually associated with Quercus species and other broad-leaved trees in humid and warm environments. The collected specimen was found on soil in a deciduous forest in October. Therefore, the preferred habitat is likely represented by soil and warm temperatures.

Inocybe latibulosa E. Horak, Matheny & Desjardin, Phytotaxa 230(3): 207, 2015 (Figs. 1 and 5)
Korean name: Bamsaek-Ttambeoseot (밤색땀버섯); derived from the color of pileus; chestnut-colored, nutbrown.
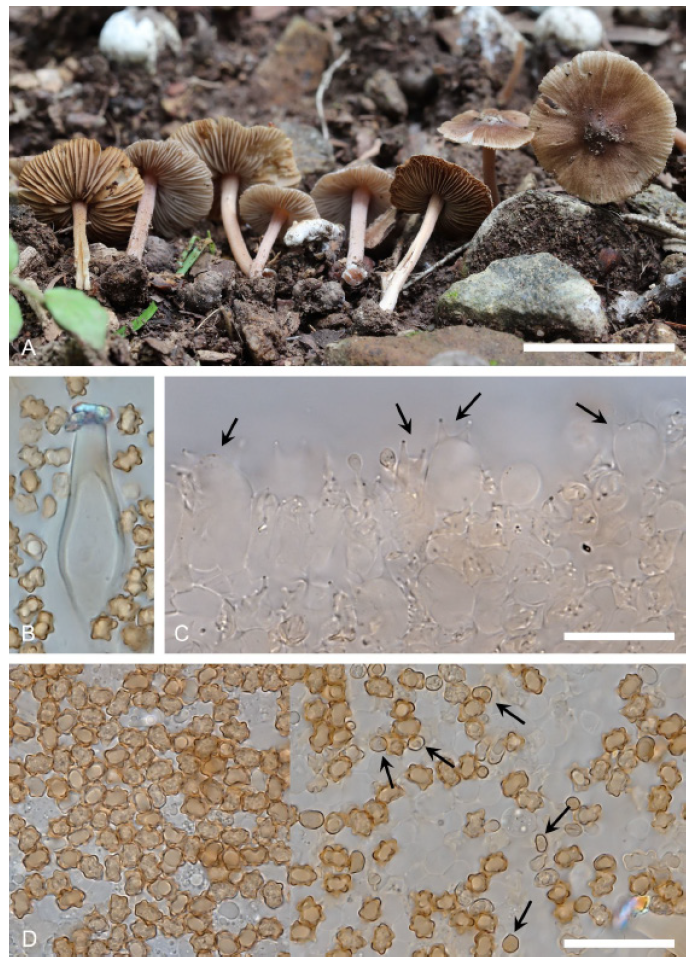
Description: Pileus 3-5 mm wide, obtusely conical, convex, or convex with a low and broad umbo, dark brown, dull, dry. Lamellae adnexed, rather distant, up to 1.5 mm wide, pale brown, fimbriate edges concolorous or whitish. Stipe 8-12×0.5-1 mm, central, cylindrical, equal, pale brown overall, subpruinose at apex, minutely fibrillose towards base, dry. Basidiospores 7.5-9×5-5.5 μm, brown, star-shaped, occasionally smooth when immature. Basidia 24-30×7-9 μm, 4-spored, clavate. Cheilocystidia 45-58×14-20 μm, subfusoid, wall thin, hyaline. Paracystidia 22-35×15-23 μm, broadly clavate or subglobose, conspicuous, hyaline. Pleurocystidia similar to cheilocystidia. Caulocystidia 35-63×7-12 μm, polymorphic, shape ranging from cylindrical to subfusoid, occasionally encrusted with brown pigment. Pileipellis short-globose or ovoid cells, 20-34 μm wide, slender fusoid, or conical. Clamp connections present.
Note: In 2015, this species was introduced as a new species in Thailand [13]. Since 2013, these specimens have been collected in Korea. The basidiospores of this species is known as smooth and amygdaliform, but Korean specimens have star-like shaped and smooth basidisopores. Some smooth basidiospores were also found when immature. In this study, six Korean specimens and two samples of I. latibulosa including type specimen are shown that they are monophyletic taxon with 100% bootstrap support values. Therefore, characteristics of basidiospores of this species is needed to confirmed with additional collections.
Inocybe stellata E. Horak, Matheny & Desjardin, Phytotaxa 230(3): 215, 2015 (Figs. 1 and 6)
Korean name: Byeol-Ttambeoseot (별땀버섯); derived from the stellatus (Lat.) star-like, shape of the basidiospores.
Description: Pileus 7-16 mm wide, dark brown, whitish fibrils or appendiculate, non-persisting veil remnants towards rimose margin. Lamellae adnexed or sinuate, ventricose, up to 3.5 mm wide; at first pale gray-brown, becoming dark brown or chocolate brown in age, edges subfimbriate, concolorous, or whitish. Stipe 18-35×1.5-2 mm, central, cylindrical, slender, equal or base slightly swollen, densely covered with conspicuous whitish, pale gray-brown or brown, with white tomentum covering the base, cortina present. Basidiospores 5-8 μm globose or subglobose, star-shaped. brown. Basidia 31-37×11.5-15 μm, 4-spored, clavate, or subcylindrical. Cheilocystidia 35-45×15-19 μm, broadly fusoid, wall thick or thin, hyaline, paracystidia 14-23×10-14 μm, clavate or vesiculose, thin-walled, hyaline. Pleurocystidia similar to cheilocystidia. Caulocystidia scattered. Pileipellis a trichoderm or cutis of cylindrical hyphae, 3.5-7 μm wide, non-gelatinized wall encrusted with brown pigment. Clamp connections present.
Note: In 2015, I. stellata was introduced as a new species [13]. The type species were collected from tropical lowland and tropical montane forests in Thailand and Bangladesh. The Korean sample was also collected during the warm and humid seasons in Korea (Table 1). Therefore, we confirmed that this species shares a similar ecological habitat. According to Horak et al. [13], nLSU ML phylogeny showed that two separate clades of I. stellata had high bootstrap values (100% and 98%, respectively). In the present study, we provided ITS phylogeny for the first time. This species was divided into three lineages, and the Korean specimen was closely related to I. stellata from Thailand (GQ893012). Morphologically, the Korean specimen has a smaller pileus and shorter stipe than I. stellata (pileus: 7-16 mm vs. 8-25 mm wide, respectively; stipe: 18-35 mm vs. 35-70 mm long, respectively). In addition, the basidiospores and basidia of the Korean specimen were smaller than those of I. stellata (basidia: 31-37×11.5-15 μm vs. 30-40×11.5-15 μm, respectively; basidiospores: 5-8 μm vs. 12-16 μm, respectively). Therefore, additional collections and multi-gene analyses are needed to elucidate these genetic variations and morphological differentiation for species identification.

Inosperma shawarense (Naseer & Khalid) Aïgnon & Naseer (Figs. 1 and 7)
Basionym: Inocybe shawarensis A. Naseer & A.N. Khalid, in Naseer, Khalid & Smith, Mycotaxon 132(4): 912, 2017.
Korean name: Gwangneung-Minttambeoseot (광릉민땀버섯); derived from the collection locality situated in South Korea; Korean genus name is newly designated as Minttambeoseot (민땀버섯), derived from the characteristics of smooth basidiospores.
Description: Pileus 30 mm in diameter, campanulate, umbonate, margin slightly incurved, surface fibrillose, rimose, dark brown at the center, becoming lighter brown in patches toward the creamy white margin. Lamellae, fimbriate, light gray when young. Stipe 2.5-4.0 cm, cylindrical, surface fibrillose, apex slightly pruinose, slightly narrower towards apex, base slightly swollen to bulbous. Basidiospores 4.5-6.2×2.7-3.5 µm, yellowish brown, smooth. Basidia 23-30×6.5-9 µm, light brown, clavate, blunt ended, 4-spored. Cheilocystidia 25-50×9.2-11 µm, clavate, thin walled. Pleurocystidia none. Pileipellis a radially orientated cutis of thin-walled hyphae, 5-7 µm in diameter, cylindrical, walls smooth or finely encrusted, light brown, blunt ends, septate. Caulocystidia at the apex, more or less similar to cheilocystidia. Stipitipellis hyphae 4-7 µm in diameter, septate, filamentous, unbranched. Clamp connections present.
Note: Inosperma shawarensis was introduced as a new species in 2017 [16]. This species belongs to the Inocybe subgen. Inosperma, which is now treated as a genus rank [6]. Recently, this species has been proposed as I. shawarense comb. nov. [17]. Most species of Inosperma bear paseoliform spores [4], and I. shawarense is characterized by a brown, fibrillose campanulate pileus, grayish lamellae, slightly bulbous stipe base, slightly pruinose silvery white stipe, small phaseoloiform spores, and clavate cheilocystidia. These morphological characteristics of Inosperma species were similar to those of the Korean collection. In addition, the Korean specimen was matched with the type species (KY616964 and KY616965) with 100% bootstrap support values (Fig. 1).

Mallocybe malenconii (R. Heim) Matheny & Esteve-Rav., in Matheny, Hobbs & Esteve-Raventós, Mycologia 112: 107, 2019 (Figs. 1 and 8)
Synonymy: Inocybe malenconii R. Heim Encyclop. Mycol., 1 Le Genre Inocybe (Paris): 163, 1931.
Inocybe malenconii var. cylindrata Takah. Kobay., Mycoscience 46(3): 184, 2005.
Korean name: Hwanggalsaeg-Yangmottambeoseot (황갈색양모땀버섯); derived from the color of pileus, light brown or yellowish brown; Korean genus name is newly designated as Yangmottambeoseot (양모땀버섯), derived from the woolly pileus.
Description: Pileus 2-2.5×0.6 cm, similar to a button mushroom when young, soon becoming plano-convex to applanate; margin inflexed at first, with a distinct cortina, expanded, somewhat bent, ochre-brown. Surface woolly-fibrillose, scurfily-squamose. Gills rather crowded, adnate, and decurrent edges slightly undulating and floccose. Stipe 3×4 cm, cylindrical, parallel, curved, concolorous with cap but paler. Basidia 30-36×8-10 µm, generally 4-spored. Basidiospores 10-13.5×4.5-6.5 µm. Cheilocystidia 30-50×14-22 µm. Stipe surface hyphae 8-12 µm wide. Cap cuticle hyphae 10-13 µm wide.
Note: In 2019, Mallocybe was newly introduced as a generic rank based on multigene molecular phylogenetic analysis [1]. It is characterized by short cheilocystidia (less than 50 µm in length). Fifty-five species of Mallocybe have been identified in the temperate regions of Africa, Asia, Australia, Europe, New Zealand, and North America. In this study, Mallocybe was recorded for the first time in Korea.

Conclusion
Among the Inocybaceae, the genus Nothocybe is only known for the type species N. distincta from India. Inocybe, Inosperma, Mallocybe, Pseudosperma, Auritella, and Tubariomyces are widespread worldwide. In this study, new records of seven species belonging to Inocybaceae, specifically Inocybe caroticolor, I. fuscidula, I. grammatoides, I. latibulosa, I. stellata, Inosperma shawarense and Mallocybe malenconii, were reported based on specimens collected in Korea. Among these, Inosperma and Mallocybe were included in the present study. In Korea, 37 species of Inocybaceae have been reported to date [7]. The most recent paper on the Inocybaceae in Korea was published in 2000 [10]. Since then, additional reports and molecular phylogenetic analyses with Korean collections have not been conducted. Therefore, it is necessary to update the traditionally recognized morphological species of Korean Inocybaceae.

