Yoosun Oh, Wonsu Cheon, Jaeduk Goh, Chang Soo Lee, and Hye Yeon Mun*
Biological Resources Research Department, Nakdonggang National Institute of Biological Resources, Sangju 37242, Korea
*Correspondence to outcastm@nnibr.re.kr
Korean Journal of Mycology (Kor J Mycol) 2025 June, Volume 53, Issue 2, pages 109-119.
https://doi.org/10.4489/kjm.2025.53.2.6
Received on April 10, 2025, Revised on June 10, 2025, Accepted on June 11, 2025, Published on June 30, 2025.
Copyright © The Korean Society of Mycology.
This is an Open Access article which is freely available under the Creative Commons Attribution-NonCommercial 4.0 International License (CC BY-NC) (https://creativecommons.org/licenses/by-nc/4.0/).
Pyrenochaetopsis is a genus of filamentous fungi within the Cucurbitariaceae family (Ascomycota, Dothideomycetes, and Pleosporales). Pyrenochaetopsis is commonly isolated from the soil, plants, and marine sediments. Species in this genus are receiving attention owing their ecological functions and biotechnological potential given their roles as plant pathogens, decomposers, and production of bioactive compounds. Five previously unrecorded Pyrenochaetopsis species were isolated from freshwater environments in Korea and identified using internal transcribed spacer (ITS) sequencing. The strains were identified as P. conf luens NNIBRFG21038, P. kuksensis NNIBRFG33605, P. leptospora NNIBRFG33604, P. microspora NNIBRFG21054, and P. uberiformis NNIBRFG33601. Their morphological and physiological characteristics, including growth patterns, spore structure, and potential antimicrobial and enzymatic activities, were examined. These findings contribute to our understanding of the Pyrenochaetopsis diversity in Korea, providing foundational data for future ecological and applied studies.
Enzyme activity, Freshwater fungi, Korea, Pyrenochaetopsis, taxonomy
Pyrenochaetopsis 속(Ascomycota, Pezizomycotina, Dothideomycetes, Pleosporales, Cucurbitariaceae)은 자낭균류에 속하는 사상성 균류로, 토양, 식물, 해양 퇴적물 등 다양한 환경에서 분리된다. 이 속은 다계통 분류군이었던 Phoma 및 Pyrenochaeta에서 형태적·분자적 차이가 있는 종들을 분리하여 재정립된 분류군이다[1]. 형태학적으로는 두꺼운 벽을 가진 피크노이드형 분생자낭(pycnidium)과 단세포 분생포자(conidia)를 형성하는 것이 특징이다[2].
최근 Pyrenochaetopsis 속은 식물병원균뿐만 아니라 식물병원균을 억제하는 길항작용을 가지는 것으로 보고되었으며, 해양 및 육상 생태계의 유기물 분해자, 그리고 천연물 생산 균주로서의 잠재적 가치가 주목받고 있다[2]. 특히 해양 및 담수와 같은 수생 환경에서의 분리 사례가 증가하고 있으며, 이들은 다양한 환경 적응 능력과 생리·생태학적 다양성을 지닌 것으로 보고되고 있다[2]. 현재까지 본 속은 전 세계적으로 24종이 보고되어 있으며[3], 국내에서는 2024년 기준 단 1종만이 기록되어 있다[4].
본 연구에서는 국내 담수 환경에서 수집한 담수 및 퇴적물로부터 균류를 분리하고 동정하였다. 그 결과, 국내에서 미기록된 Pyrenochaetopsis 속 5종을 확인하였으며, 이들 균주의 형태학적 특징, 분자 계통학적 분석, 그리고 기초적인 생리활성 정보를 보고하고자 한다.
담수균류를 분리하기 위하여 경기도 용인시와 전라북도 진안군의 담수 환경에서 담수 시료와 퇴적물(물 속에 잠긴 토양)을 채집하였다. 담수 시료는 조사 현장에서 핸드펌프와 니트로셀룰로오스 멤브레인 필터(nitrocellulose membrane filter, pore size 0.45 µm; MCE membrane, MFMillipore™, Burlington, MA, USA)를 이용하여 여과하였다. 시료가 여과된 필터 면은 스트렙토마이신 100 ppm이 첨가된 워터 배지(water agar, WA; 20 g/L agar)에 부착한 뒤, 15℃에서 1일간 배양하였다. 이후 멤브레인 필터를 제거하고, 실체현미경을 이용하여 발아한 포자를 분리하여 V8 배지(V8 agar; 8% V8 주스 [v/v], 1.5% agar [w/v], pH 6.0으로 10 N NaOH로 조정)에 접종하여 배양하였다. 담수 환경에서 채집한 퇴적물 시료는 10-³로 희석한 후, 스트렙토마이신 100 ppm이 첨가된 감자한천 배지(potato dextrose agar, PDA; Difco, Detroit, MI, USA)에 도말하여 15℃에서 3일간 배양하였다. 배양된 균주들은 단포자 분리를 통해 V8A 배지에서 순수 분리하였으며, 확보된 균주의 균체는 PDA 배지(3.9% potato dextrose agar powder [w/v]; Difco, Sparks, MD, USA)에서 계대배양하여 확보하였다.
생육 환경에 따른 특성을 조사하기 위하여 malt extract agar (MEA; 2% malt extract [w/v], 2% agar [w/v]), oatmeal agar (OA; 7.25% oatmeal agar powder [w/v]; Difco, Sparks, MD, USA), PDA의 세 가지 배지를 실험에 사용하였다. 분리된 균류의 포자 관찰 및 이미지 확보는 광학 현미경(H550S; Nikon, Tokyo, Japan)을 이용하여 수행하였다.
분리 균주의 동정을 위해 ITS (internal transcribed spacer) region [ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) 및 ITS4 (5′-TCCTCCGCTTATTGATATGC-3′)] 및 LSU (large subunit) [LR0R (5′-ACCCGCTGAACTTAAGC-3′) 및 LR7 (5′-TACTACCACCAAGATCT-3′)]의 염기서열을 사용하였으며[5,6], BLAST(Basic Local Alignment Search Tool)를 통해 데이터베이스에 등록된 균류들과 상동성을 비교하였다(Table 1). 이후 MEGA 11 프로그램[7]을 이용하여 계통수를 작성하였다.
국내 미기록종의 산업적 활용 가능성을 평가하기 위하여 항균 활성 및 효소 활성을 기초적으로 검정하였다. 고추시들음병원균 Fusarium solani와 고추역병균 Phytophthora capsici, 그리고 실험 균주의 아가 플러그를 PDA 배지 상에서 대치배양하였다. 아밀라아제, 글리코시다아제, 라카아제, 키티나아제 활성을 검정하기 위해 각각 다음과 같은 방법를 사용하였다: 아밀라아제 분석을 위해 α-Amylase Assay Kit (Megazyme, Bray, Ireland), 글리코시다아제 분석을 위해 β-Glucosidase Assay Kit (MAK129, Sigma-Aldrich, St. Louis, USA), 키티나아제 분석을 위해 Chitinase Assay Kit (CS0980, Sigma-Aldrich, St. Louis, USA), 락카아제 분석을 위해 More 등[8]의 방법을 실험에 사용하였다.
Table 1. Accession number of Pyrenochaetopsis.
테이블
ITS, internal transcribed spacer; LSU, large subunit.
균학적 특성: 고체 배지 상에서 25℃에서 7일간 배양하였을 때, 콜로니는 10–15 mm 정도로 느리게 생장하였다. MEA 배지에서는 앞면이 크림색을 띠었고, 뒷면은 연한 황갈색을 거쳐 가장자리가 다시 크림색으로 변하였다. OA 배지에서는 앞면이 연한 회갈색이며 가장자리는 크림색, 뒷면은 밝은 베이지색을 나타냈다. PDA 배지에서는 앞면이 짙은 올리브색에서 회녹색을 띠었고, 뒷면은 회녹색이며 가장자리는 크림색이었다. 콜로니는 밀도가 높고 균일하게 확산되었으며, 둥근 형태를 보였다. 단단한 형태의 분생자낭이 형성되었으며, 포자는 끝이 둥근 막대형(cylindrical)으로 관찰되었고, 크기는 약 5–6 µm × 1–1.5 µm였다(Fig. 2, Table 2).
분리원: 담수퇴적물
표본 정보: 경기도 용인시 처인구, 2019.03.07., NNIBRFG21038(FBCC-F1371), 국립낙동강생물자원관, ITS: PV368328, LSU: PV715656
비고: NNIBRFG21038 균주는 ITS 영역의 염기서열을 BLAST를 이용해 분석한 결과, P. confluens UTHSC DI16-303 균주와 100% (516/516, NR160338) 상동성을 나타냈다. 계통수 상에서도 NNIBRFG21038은 P. confluens CBS142459 균주와 같은 clade에 속하는 것으로 확인되어, 최종적으로 P. confluens로 동정되었다(Fig. 1). 한편, P. confluens는 2018년에 처음 보고된 종으로 인간의 혈액 시료에서 분리된 것으로 알려져 있다[9].
본 균주는 Fusarium solani 및 Phytophthora capsici에 대해 강한 항균활성을 나타냈으며, 효소활성 중에서는 락카아제와 키티나아제가 약하게, 아밀라아제는 매우 강하게 나타났다. 또한, PDB에 액체배양 시 오렌지빛이 도는 노란색 색소를 생성하였다(Table 3).
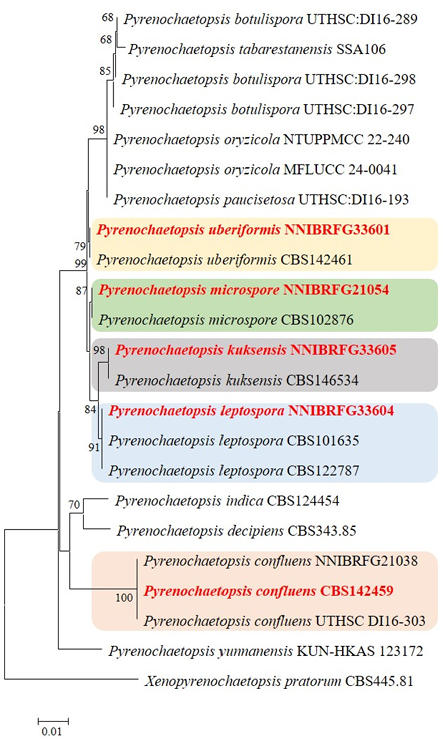
Fig. 1. Phylogenetic tree of unrecorded Pyrenochaetopsis species isolated from freshwater environments and related taxa, based on neighbor-joining analysis of internal transcribed spacer (ITS) and large subunit (LSU) region sequences using MEGA 11 [7]. Bootstrap values (1,000 replicates) are shown next to the branches. Values less than 50% were deleted. Evolutionary distances were computed using the Kimura 2-parameter method. Xenopyrenochaetopsis pratorum CBS445.91 was used as the outgroup. Unrecorded strains are highlighted in bold and red.

Fig. 2. Morphological characters of P. confluens NNIBRFG21038 (FBCC-F1371). A: Mycelial growth on MEA, OA and PDA for 7 days at 25℃. B: Pycnidia ×10. C: Conidiophore and D: Conidia ×400, scale bar = 10 µm. MEA, malt extract agar; OA, oatmeal agar; PDA, potato dextrose agar.
Table 2. Morphological characteristics of Pyrenochaetopsis.
테이블
MEA, malt extract agar; OA, oatmeal agar.
Table 3. Physiological characteristics of five Pyrenochaetopsis species isolated from freshwater environments in Korea.
테이블
FBCC: Freshwater Bioresources Culture Collection. *E: not examined; -: no activity; +: slightly activity; ++: moderate activity; +++: strong activity.
균학적 특성: 고체 배지 상에서 25℃에서 7일간 배양하였을 때, 콜로니는 20–25 mm 정도로 느리게 생장하였으며, OA 배지에서 가장 밀도 높게 자랐다. MEA 배지에서는 앞면이 회백색을 띠고, 뒷면은 옅은 갈색이었다. OA 배지에서는 콜로니의 앞면이 연한 회백색을, 뒷면은 크림색을 나타냈다. PDA 배지에서는 앞면과 뒷면 모두 회녹색을 띠었으며, 가장자리는 크림색으로 나타났다. 콜로니는 대부분 중심부가 약간 융기된 형태를 보였다. 단단한 형태의 분생자낭이 형성되었으며, 포자는 끝이 둥근 막대형(ovoid to cylindrical)으로 관찰되었고, 크기는 약 2–3 µm × 0.5–1 µm였다(Fig. 3, Table 2).
분리원: 담수퇴적물
표본 정보: 전라북도 진안군 마령면, 2021.04.01., NNIBRFG33605(FBCC-F1671), 국립낙동강생물자원관, ITS: PV368329, LSU: PV715657
비고: NNIBRFG33605 균주는 ITS 영역의 염기서열을 BLAST를 이용해 분석한 결과, Pyrenochaetopsis sp. MAFF307189 균주와 100% (429/429, LC433836) 상동성을 나타냈으며 계통수 상에서 NNIBRFG33605는 P. kuksensis CBS146534 균주와 같은 clade에 속하는 것으로 확인되어, 최종적으로 P. kuksensis로 동정되었다(Fig. 1). 한편, P. kukensis는 부식된 나무조각에서 분리된 것으로 보고되었으며, 비교적 최근에 보고된 종이다[10].
본 균주는 PDB에 액체배양 시 밝은 주황빛 노란색 색소를 생성하고, 항균활성, 인산가용화능 및 효소활성 중 키티나아제와 아밀라아제가 약하게 있는 것을 확인하였다 (Table 3).
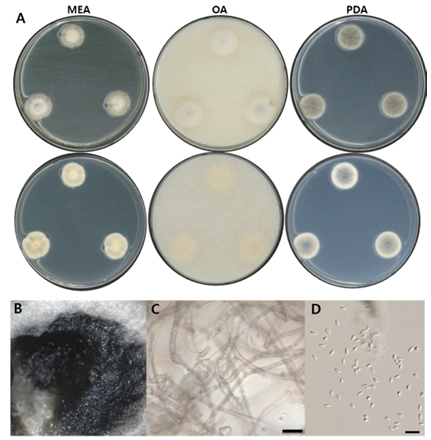
Fig. 3. Morphological characters of P. kuksensis NNIBRFG33605 (FBCC-F1671). A: Mycelial growth on MEA, OA and PDA for 7 days at 25℃. B: Pycnidia ×10. C: Conidiophore and D: conidia, ×400, scale bar = 10 µm. MEA, malt extract agar; OA, oatmeal agar; PDA, potato dextrose agar.
균학적 특성: 고체 배지 상에서 25℃에서 7일간 배양하였을 때, 콜로니는 10–15 mm 정도로 느리게 생장하였으며, OA 배지에서 생장률이 가장 높았다. MEA 배지에서는 콜로니가 크림색에서 회녹색을 띠다가, 가장자리는 다시 크림색을 나타냈다. OA 배지에서는 앞면이 옅은 회갈색을 띠며, 가장자리는 흐릿한 형태로 확산되었고, 뒷면은 연한 크림색을 띠었다. PDA 배지에서는 콜로니 중심부가 밀도가 높고 융기된 형태로 크림색을 띠었으며, 바깥쪽은 진한 회색을 띠다가 가장자리에 크림색 띠가 형성되었다. 단단한 형태의 분생자낭이 형성되었으며, 포자는 끝이 둥근 막대형(cylindrical)으로 관찰되었고, 크기는 5–6 µm × 1–1.5 µm였다(Fig. 4, Table 2).
분리원: 담수퇴적물
표본 정보: 전라북도 진안군 마령면, 2021.04.01., NNIBRFG33604(FBCC-F1672), 국립낙동강생물자원관, ITS: PV368330, LSU: PV715658
비고: NNIBRFG33604 균주는 ITS 영역의 염기서열을 BLAST를 이용해 분석한 결과, P. leptospora CBS101635 균주와 100% (534/534, MF795793) 상동성을 나타냈다. 계통수 상에서도 NNIBRFG33604는 P. leptospora CBS101635 및 CBS122787 균주와 같은 clade에 속하는 것으로 확인되어, 최종적으로 P. leptospora로 동정되었다(Fig. 1). P. leptospora는 Pyrenochaeta leptospora 였다가 2010년 재분류되어 현재의 종명으로 변경되었으며, 잔디의 잎이나, 밀밭의 토양 등에서 분리되었다[2].
본 균주는 인산가용화능과 Fusarium solani 및 Phytophthora capsici에 대해 항균활성을 나타냈으며, PDB에 액체배양 시 진한 노란색 계열의 색소를 생성하고, 효소활성 중 키티나아제와 아밀라아제가 약하게 있는 것을 확인하였다 (Table 3).
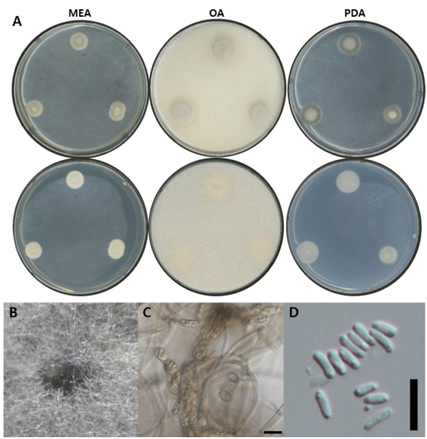
Fig. 4. Morphological characters of P. leptospora NNIBRFG33604 (FBCC-F1672). A: Mycelial growth on MEA, OA and PDA for 7 days at 25℃. B: Pycnidia ×10. C: Conidiophore and D: conidia, ×400, scale bar = 10 µm. MEA, malt extract agar; OA, oatmeal agar; PDA, potato dextrose agar.
균학적 특성: 고체 배지 상에서 25℃에서 7일간 배양하였을 때, 콜로니는 약 15–20 mm 정도로 느리게 생장하였으며, OA 배지에서 생장률이 가장 높았다. MEA 배지에서는 앞면과 뒷면 모두 크림색을 나타냈다. OA 배지에서는 콜로니가 연한 회색을 띠었고, 가장자리는 흐릿한 형태의 크림색을 보였으며, 뒷면 전체는 크림색을 띠었다. PDA 배지에서는 다소 두터운 콜로니가 형성되었고, 앞면은 회녹색을 띠다가 가장자리는 크림색으로 생장하였다. 뒷면은 옅은 회색을 띠었으며, 가장자리 부분에서는 크림색을 나타냈다. 단단한 형태의 분생자낭이 형성되었고, 포자는 끝이 둥근 막대형(ovoid to cylindrical)으로 관찰되었으며, 크기는 약 2–3 µm × 0.5–1 µm였다(Fig. 5, Table 2).
분리원: 담수
표본 정보: 경기도 용인시 처인구, 2019.03.07., NNIBRFG21054(FBCC-F1368), 국립낙동강생물자원관, ITS: PV368331, LSU: PV715659
비고: NNIBRFG21054 균주는 ITS 영역의 염기서열을 BLAST를 이용해 분석한 결과, P. microspora CBS102876 균주와 99% (512/514, NR160059) 상동성을 나타냈다. 계통수 상에서도 NNIBRFG21054는 P. microspora CBS102876 균주와 같은 clade에 속하는 것으로 확인되어, 최종적으로 P. microspora로 동정되었다(Fig. 1). P. microspora는 이전에는 Phoma leveillei var. microspora 로 분류되었으나, 2010년에 새롭게 재정립 종으로 Skadar 호수의 담수, 커피나무 잎 등에서 분리된 것으로 보고되었다[2].
NNIBRFG21054는 Fusarium solani 및 Phytophthora capsici에 대해 항균활성 및 인산가용화능을 나타냈으며, 색소는 생성하지 않지만 락카아제, 키티나아제, 아밀라아제 효소활성이 약하게 나타나는 것을 확인하였다 (Table 3).
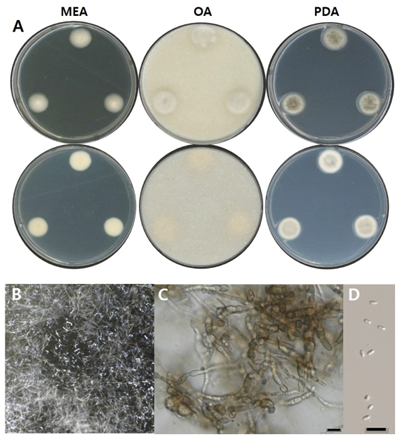
Fig. 5. Morphological characters of P. microspora NNIBRFG21054 (FBCC-F1368). A: Mycelial growth on MEA, OA and PDA for 7 days at 25℃. B: Pycnidia ×10. C: Conidiophore and D: conidia, ×400, scale bar = 10 µm. MEA, malt extract agar; OA, oatmeal agar; PDA, potato dextrose agar.
균학적 특성: 고체 배지 상에서 25℃에서 7일간 배양하였을 때, MEA 배지에서는 콜로니가 5–10 mm 정도로 매우 느리게 생장하였으며, OA 및 PDA 배지에서는 15–20 mm 정도로 생장하였다. MEA 배지에서는 앞면과 뒷면 모두 크림색을 나타냈다. OA 배지에서는 콜로니가 다소 두껍고 밀도 높게 생장하였으며, 앞면은 전체적으로 연한 회색을 띠고, 가운데와 가장자리에서는 연한 회녹색이 나타났다. 뒷면은 크림색이었다. PDA 배지에서는 앞면 중심부가 회녹색을 띠고 가장자리는 크림색이었으며, 뒷면은 중심부에서 연한 회색을 띠다가 전체적으로 크림색으로 변화하였다. 단단한 형태의 분생자낭이 형성되었고, 포자는 끝이 둥근 막대형(cylindrical)으로 관찰되었으며, 크기는 약 3–4 µm × 1–1.5 µm였다(Fig. 6, Table 2).
분리원: 담수퇴적물
표본 정보: 전라북도 진안군 마령면, 2021.04.01., NNIBRFG33601(FBCC-F1674), 국립낙동강생물자원관, ITS: PV368332, LSU: PV715660
비고: NNIBRFG33601 균주는 ITS 영역의 염기서열을 BLAST를 이용해 분석한 결과, P. uberiformis RZ-6a 균주와 99% (531/533, OR072742) 상동성을 나타냈으며 계통수 상에서 NNIBRFG33601은 P. uberiformis CBS142461 균주와 같은 clade에 속하는 것으로 확인되어, 최종적으로 P. uberiformis로 동정되었다(Fig. 1). P. uberiformis는 2018년에 인간의 귀 병변에서 최초 분리 된 것으로 보고되었으나[9], 본 연구에서는 국내 하천 퇴적토에서 분리되었다.
NNIBRFG33601은 Fusarium solani 및 Phytophthora capsici에 대해 항균활성, 인산가용화 및 효소활성 중 키티나아제, 글리코시다아제와 아밀라아제가 모두 약하게 나타나며, 색소는 생성하지 않았다(Table 3).
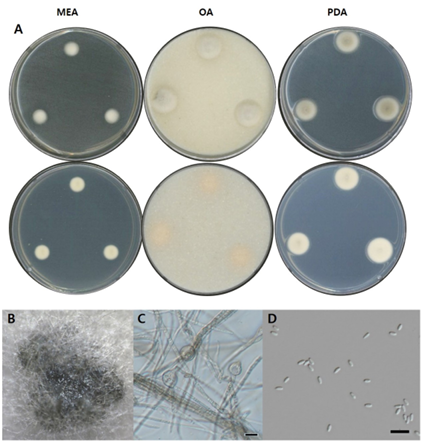
Fig. 6. Morphological characters of P. uberiformis NNIBRFG33601 (FBCC-F1674). A: Mycelial growth on MEA, OA and PDA for 7 days at 25℃. B: Pycnidia, ×10. C: Conidiophore and D: Conidia, ×400, scale bar = 10 µm. MEA, malt extract agar; OA, oatmeal agar; PDA, potato dextrose agar.
본 연구에서는 국내 담수 환경에서 수집한 시료로부터 균류를 분리하고 동정한 결과, 국내 미기록 Pyrenochaetopsis 속 5종을 확인하였다. 확인된 균주는 각각 P. confluens, P. kuksensis, P. leptospora, P. microspora, P. uberiformis이며, ITS 염기서열 분석을 통해 동정되었다. 분리 균주의 형태적 특징 및 생장 특성을 배지별로 비교하였으며, 항균 활성 및 효소 활성 등 기초적인 생리활성을 평가하였다. 이 연구는 국내 담수 환경에서의 Pyrenochaetopsis 속의 다양성 이해를 넓히고 생리생태적 기초자료를 제공한다.
The authors declare no competing interests.
This work was supported by a grant from the Nakdonggang National Institute of Biological Resources (NNIBR)(NNIBR20251105) and the Korea Environment Industry & Technology Institute (KEITI) through a project to make multi-ministerial national biological research resources a more advanced program (2021003420002) funded by the Korea Ministry of Environment (MOE).
1. Magaña-Dueñas V, Stchigel AM, Cano-Lira JF. New coelomycetous fungi from freshwater in Spain. J Fungi 2021;7:368. [DOI]
2. de Gruyter J, Woudenberg JHC, Aveskamp MM, Verkley GJM, Groenewald JZ, Crous PW. Systematic reappraisal of species in Phoma section Paraphoma, Pyrenochaeta and Pleurophoma. Mycologia 2010;102:1066-81. [DOI]
3. Catalogue of Life. Fungi [Internet]. Amsterdam: Catalogue of Life; 2025 [cited 2025 April 10]. Available from https://www.catalogueoflife.org/index?taxonKey=F.
4. National Institute of Biological Resources, Korean Society of Mycology, Microbiological Society of Korea. National species list of Korea. Incheon: National Institute of Biological Resources; 2024.
5. White TJ, Bruns TD, Lee SB, Taylor JW. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, editors. PCR protocols: A guide to methods and applications. New York: Academic Press; 1990. p. 315-22. [DOI]
6. Vilgalys R, Hester M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J Bacteriol 1990;172:4238-46. [DOI]
7. Tamura K, Stecher G, Kumar S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol Biol Evol 2021;38:3022-7. https://doi.org/10.1093/molbev/msab120 [DOI]
8. More SS, Renuka PS, Pruthvi K, Swetha M, Malini S, Veena SM. Isolation, purification, and characterization of fungal laccase from Pleurotus sp. Enzyme Res 2011;2011:248735. https:// doi.org/10.4061/2011/248735 [DOI]
9. Valenzuela-Lopez N, Cano-Lira JF, Guarro J, Sutton DA, Wiederhold N, Crous PW, Stchigel AM. Coelomycetous Dothideomycetes with emphasis on the families Cucurbitariaceae and Didymellaceae. Stud Mycol 2018;90:1-69. [DOI]
10. Špetík M, Berraf-Tebbal A, Pokluda R, Eichmeier A. Pyrenochaetopsis kuksensis (Pyrenochaetopsidaceae), a new species associated with an ornamental boxwood in the Czech Republic. Phytotaxa 2021;498:177-85. [DOI]