¹Integrated Science Education, Graduate School, Korea National University of Education, Cheongju-si, Chungbuk 28173, Korea
²Department of Biology Education, Korea National University of Education, Cheongju-si, Chungbuk 28173, Korea
*Correspondence to eomah@knue.ac.kr
Korean Journal of Mycology (Kor J Mycol) 2025 June, Volume 53, Issue 2, pages 137-144.
https://doi.org/10.4489/kjm.2025.53.2.9
Received on May 27, 2025, Revised on June 18, 2025, Accepted on June 19, 2025, Published on June 30, 2025.
Copyright © The Korean Society of Mycology.
This is an Open Access article which is freely available under the Creative Commons Attribution-NonCommercial 4.0 International License (CC BY-NC) (https://creativecommons.org/licenses/by-nc/4.0/).
Prunus × yedoensis, which is widely cultivated for ornamental purposes, is highly susceptible to witches’ broom disease caused by Taphrina wiesneri. Conventional physical and chemical control measures have certain limitations due to environmental concerns, thereby highlighting the need for alternative approaches. In this study, we sought to isolate and identify endophytic fungi from Prunus × yedoensis and evaluate their potential as biological control agents against T. wiesneri using dual culture assays. A total of 45 endophytic fungal species were isolated and identified based on an analysis of internal transcribed spacer sequences. Fungal species composition was found to differ between leaves and twigs, with Diaporthe eres and Nothophoma quercina being the most prevalent species. Dual culture assays revealed that Aspergillus flavus, and Trichoderma guizhouense have significant inhibitory effects on T. wiesneri (inhibition index > 0.5). These findings indicate that endophytic fungi could be harnessed to facilitate the suppression of witches’ broom disease in Prunus × yedoensis
Biological Control Agent, Dual Culture, Endophytic fungi, Prunus × yedoensis, Taphrina wiesneri, Witches’ broom
Prunus × yedoensis Matsum., a plant prized for its ornamental value, is widely planted in urban landscapes and public parks [1]. However, its susceptibility to witches’ broom disease caused by the fungus Taphrina wiesneri poses a serious threat to its health and longevity [2]. Infected twigs tend to be characterized by abnormal shoot proliferation and leaf expansion, leading to suppressed flowering and reduced tree vitality. In severe cases, twig dieback occurs within a few years, and prolonged infection can result in tree mortality [3,4].
Taphrina wiesneri has been established to colonize host tissues and produce phytohormones, such as indole-3-acetic acid and cytokinin, that contribute to disrupting the host’s hormonal balance and inducing disease symptoms [5,6]. Current control strategies include the physical removal of infected twigs and the application of triazole fungicides, such as tebuconazole and difenoconazole [7]. However, these approaches are often insufficient for long-term disease management and raise ecological concerns relating to chemical use and pathogen resistance [8,9].
Endophytic fungi, which reside within plant tissues without causing disease, have garnered attention for their potential as biological control agents [10,11]. These fungi can inhibit plant pathogens via competition, mycoparasitism, or the production of antifungal compounds, and the findings of several studies have provided evidence of the antagonistic activity of endophytes against important phytopathogens, thereby highlighting their potential utility in sustainable plant disease management [12–14].
In this study, we sought to isolate and identify endophytic fungi from healthy Prunus × yedoensis tissues and to evaluate their antagonistic activity against T. wiesneri using dual culture assays. By identifying potential biocontrol candidates, our findings in this study will provide a basis for developing eco-friendly alternatives for managing witches’ broom disease in cherry trees.
In April 2024, samples of healthy leaves and twigs were collected from 12 Prunus × yedoensis trees in Seoul, and in September 2024, from five trees in Cheongju. Having initially washed under running tap water, these samples were sequentially surface-sterilized using 35% hydrogen peroxide (H₂O₂) for 2 min, followed by 70% ethanol for 30 s, and were then rinsed three times with sterile water [15]. The surfacesterilized tissues were thereafter cut into 10 × 10 mm (leaves) or 10 mm (twigs) segments and placed on potato dextrose agar (PDA; Kisan Bio, Seoul, Korea). Plates were incubated at 25°C and monitored for the emergence of fungal colonies, which were subsequently sub-cultured onto fresh PDA to obtain pure isolates.
Genomic DNA was extracted from the isolates using a HiGene™ Genomic DNA Prep Kit for Fungi (BioFACT, Korea). The internal transcribed spacer (ITS) region, including ITS1, 5.8S rDNA, and ITS2 sequences, was amplified using the primer pair ITS1-F and ITS4 [16,17]. Additional DNA regions, namely, the large subunit (LSU) and translation elongation factor-1-α (Tef1α), were also amplified, using LR0R/ LR5 [18] and EF1-983F/EF1-1567R [19], respectively. Sequences were identified on the basis of BLAST (Basic Local Alignment Search Tool) searches against the NCBI (National Center for Biotechnology Information) GenBank database, and have been submitted to this database.
Dual culture assays were performed to assess the antagonistic activity of endophytic fungal isolates against Taphrina wiesneri strain KACC45487, which was obtained from the Korean Agricultural Culture Collection (KACC). A 7-mm-diameter agar plug of T. wiesneri was placed 10 mm from the edge of 90-mm-diameter PDA plates and incubated at 25°C for 7 days. Subsequently, a plug of an endophytic fungal isolate was placed on the opposite side of plates, and the plates were incubated at 25°C for a further 21 days. Each assay was performed in triplicate. As a control, T. wiesneri was cultured alone under the same conditions. The area of T. wiesneri colonies was measured using ImageJ software [20], and the inhibition index (I) was calculated using the following formula:
I = (A control – A treatment )/A control
where A control is the colony area of T. wiesneri in control plates, and A treatment is the colony area in dualculture plates [21]. Statistical analysis of the inhibitory effects was conducted using Student’s t-test in R (v. 4.2.2), for which, p-values < 0.05 were considered significant.
A total of 204 fungal strains were isolated from the leaves and twigs of Prunus × yedoensis. On the basis of BLAST analysis of the ITS sequences of these isolates against accessions in the GenBank database, strains with ≥ 98% similarity were identified at the species level, whereas those with lower similarity were assigned to the genus level (Table 1). A total of 45 endophytic fungal species from 21 genera were identified from Prunus × yedoensis, among which 33 species from 19 genera were isolated from leaves, and 25 species from nine genera were obtained from twigs. Common species found in both organs included Alternaria alternata, Aureobasidium pullulans, and Colletotrichum fioriniae (Table 1). Nothophoma quercina was identified as the most frequently isolated species from leaves (41.2%), followed by Paraconiothyrium brasiliense (35.3%) and Dothiorella gregaria (29.4%), whereas Diaporthe eres was the most common species detected in twigs (70.6%), followed by Botryosphaeria dothidea (52.9%) and Diaporthe nobilis (41.2%) (Table 1). Statistical analysis of species diversity indices revealed that values of the ShannonWiener diversity index were significantly higher for fungi in twigs than those in leaves (p < 0.05) (Fig. 1). However, we detected no significant differences between leaves and twigs with respect to species richness and evenness. These findings accordingly reveal differences in the composition and diversity of endophytic fungi in the leaves and twigs of Prunus × yedoensis, which is consistent with previously reported findings indicating that fungal communities differ depending on plant tissue and environmental conditions [21,22].
Table 1. Frequency of endophytic fungi isolated from the leaves and twigs of Prunus × yedoensis
테이블
ITS, internal transcribed spacer.
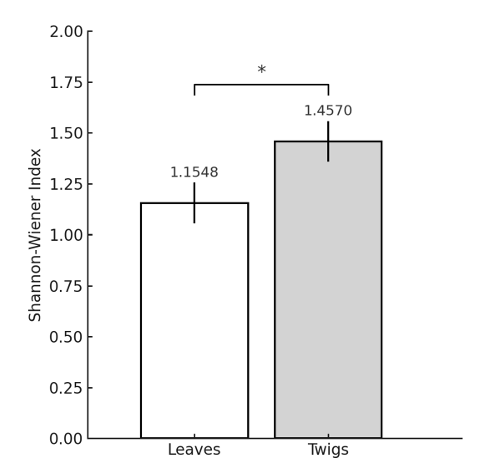
Fig. 1. Comparison of the Shannon–Wiener diversity index values obtained for endophytic fungi isolated from the leaves and twigs of Prunus × yedoensis. Values of the Shannon–Wiener diversity index for twig isolates were approximately 0.138- to 0.464-fold higher than those for leaf isolates (t = -2.614, p = 0.016). * Indicates p < 0.05.
Twelve endophytic fungal species, selected based on their prevalence in both sampling regions and plant organs, were tested against T. wiesneri KACC45487 using dual culture assays. The results revealed that although none of the assessed endophytic fungi produced zones of inhibition of T. wiesneri growth, within 7 days, some fungal isolates had overgrown the pathogen colony. Among these, five isolates were found to have caused a significant suppression of pathogen growth (p < 0.05) (Fig. 2), with Aspergillus flavus 24N0267 and Trichoderma guizhouense 24N0293 isolated from leaves being characterized by inhibition indices (I) exceeding 0.5 (Table 2).
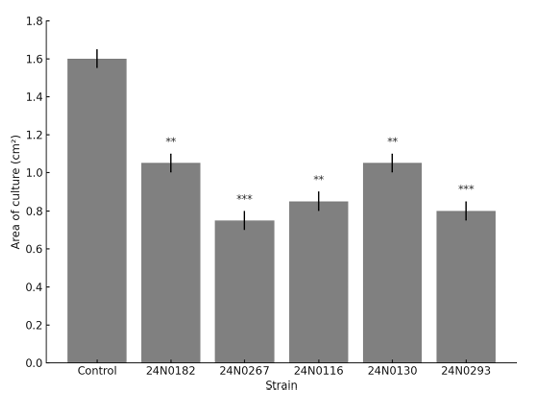
Fig. 2. Comparison of the culture area of Taphrina wiesneri KACC45487 measured on the 21st day of dual culture with selected endophytic fungi. Bars represent mean areas with standard errors. Asterisks indicate significant differences compared with the control (KACC45487) based on analysis using Student’s t-test: ** p <0.01, *** p < 0.001.
Table 2. Molecular identification and the inhibition index of five endophytic fungal strains against Taphrina wiesneri KACC45487
테이블
ITS, internal transcribed spacer; LSU, large subunit; Tef1α, translation elongation factor-1.
Aspergillus flavus is widely encountered as an endophytic fungus in woody and herbaceous plants worldwide [23,24]. It produces mycotoxins, such as aflatoxin B1 (AFB1) and aspergillic acid, along with extracellular hydrolytic enzymes, including pectinase and protease, which contribute to fungal defense mechanisms [25,26]. These enzymes can potentially degrade the cell walls of other fungi, thereby contributing to antagonistic activity of this species. However, AFB1 has been established to be a potent carcinogen that contaminates crops such as peanuts and corn, and thus further studies are necessary to assess ecological safety of A. flavus for biocontrol applications [27]. Similarly, Alternaria alternata produces AAL- and AF-toxins and can act as an opportunistic pathogen in several crops [28,29]. Moreover, Fusarium verticillioides induces wilt and rot in maize [30]. Therefore, additional studies are required to reduce the toxicity and enhance the stability of these fungi for use as biocontrol agents.
In contrast, Pestalotiopsis microspora has been established to produce pestacin, an antifungal compound, and taxol, an anticancer agent [31,32], whereas Trichoderma guizhouense is an efficient producer of cellulase and has been applied to enhance crop productivity [33]. It is accordingly speculated that its potential utility as a biocontrol agent may involve mycoparasitism mediated via the production of cell walldegrading enzymes. On the basis of the evidence obtained to data, these two species are thus considered promising candidates for safe and effective biological control applications [34].
Given that T. wiesneri resides within host tissues and induces disease symptoms, employing endophytic fungi that naturally inhabit the same niche without harming the plant represents a sustainable and ecologically safe strategy for disease management. If further experiments confirm the inhibitory effects of selected endophytic fungi on T. wiesneri in vivo, this approach could be practically applied in disease control. Moreover, if the five fungal species identified in this study are found at significantly lower frequencies in diseased trees than in healthy ones, this would provide additional evidence in support of their role as potential biocontrol agents against witches’ broom disease.
In this study, we identified 45 endophytic fungal species isolated from Prunus × yedoensis and assessed their potential as biocontrol agents against T. wiesneri. Among these fungal isolates, five strains were demonstrated to have significant inhibitory effects against T. wiesneri in dual culture assays. Further studies should evaluate the field efficacy and ecological safety of these strains to facilitate the development of sustainable biological control strategies against witches’ broom disease.
The authors declare no conflicts of interest.
1. Shi S, Li J, Sun J, Yu J, Zhou S. Phylogeny and classification of Prunus sensu lato (Rosaceae). J Integr Plant Biol 2013;55:1069-79. [DOI]
2. Kim KH. Pest and disease control – Witches’ broom disease of cherry trees. Landsc Trees 2014;139:49-51.
3. Byun BH. Plant diseases and pests in January and February – Witches’ broom disease of cherry trees. Landsc Trees 1997;36:30.
4. Komatsu M, Taniguchi M, Matsushita N, Takahashi Y, Hogetsu T. Overwintering of Taphrina wiesneri within cherry shoots monitored with species-specific PCR. J Gen Plant Pathol 2010;76:363-9. [DOI]
5. Johnston JC, Trione EJ. Cytokinin production by the fungi Taphrina cerasi and T. deformans. Can J Bot 1974;52:1583-9. [DOI]
6. Masuya H, Kikuchi T, Sahashi N. New insights to develop studies on Witch’s broom caused by Taphrina wiesneri. J Jpn For Soc 2015;97:153-7. [DOI]
7. Seo ST, Kim KH, Shin CH, Lee SH, Kim YM, Park JH, Shin SC. Control efficacy of fungicides on cherry witches’ broom caused by Taphrina wiesneri. Res Plant Dis 2009;15:13-6. [DOI]
8. Guthman J. Wilted: Pathogens, chemicals, and the fragile future of the strawberry industry. Berkeley, CA: University of California Press; 2019. [DOI]
9. Son SY, Lee SK, Seo ST. Detection of overwintering sites inhabited by cherry witches’ broom pathogen Taphrina wiesneri with species-specific PCR in Korea. J Korean For Soc 2015;104:332-5. [DOI]
10. Hosseyni-Moghaddam MS, Soltani J. Bioactivity of endophytic Trichoderma fungal species from the plant family Cupressaceae. Ann Microbiol 2014;64:753-61. [DOI]
11. Tiwari P, Bae H. Endophytic fungi: Key insights, emerging prospects, and challenges in natural product drug discovery. Microorganisms 2022;10:360. [DOI]
12. Nzabanita C, Zhang L, Wang Y, Wang S, Guo L. The wheat endophyte Epicoccum layuense J4-3 inhibits Fusarium graminearum and enhances plant growth. J Fungi 2023;10:10. [DOI]
13. Zhang J, Bayram Akcapinar G, Atanasova L, Rahimi MJ, Przylucka A, Yang D, Kubicek CP, Zhang R, Shen Q, Druzhinina IS. The neutral metallopeptidase NMP1 of Trichoderma guizhouense is required for mycotrophy and self-defense. Environ Microbiol 2016;18:580-97. [DOI]
14. Xu Y, Zhang J, Shao J, Feng H, Zhang R, Shen Q. Extracellular proteins of Trichoderma guizhouense elicit an immune response in maize (Zea mays) plants. Plant Soil 2020;449:13349. [DOI]
15. Kim JH, Kim DY, Park H, Cho JH, Eom AH. Neocosmospora rubicola, an unrecorded endophytic fungus isolated from roots of Glycyrrhiza uralensis in Korea. Kor J Mycol 2017;45:63-7. [DOI]
16. Gardes M, Bruns TD. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol Ecol 1993;2:113-8. [DOI]
17. White TJ, Bruns TD, Lee SB, Taylor JW. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, editors. PCR protocols: A guide to methods and applications. New York: Academic Press; 1990. p. 315-22. [DOI]
18. Vilgalys R, Hester M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J Bacteriol 1990;172:4238-46. [DOI]
19. Rehner S. Primers for elongation factor 1-a (EF1-a) [Interent]. Beltsville, MD: Insect Biocontrol Laboratory; 2001 [Accessed 2025 Apr 30]; Available from: https://web.archive.org/ web/20060518132929/http://ocid.NACSE.ORG/research/deephyphae/EF1primer.pdf.
20. Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nat Methods 2012;9:671-5. [DOI]
21. Al-Reza SM, Rahman A, Ahmed Y, Kang SC. Inhibition of plant pathogens in vitro and in vivo with essential oil and organic extracts of Cestrum nocturnum L. Pestic Biochem Physiol 2010;96:86-92. [DOI]
22. Sudheep NM, Sridhar KR. Non-mycorrhizal fungal endophytes in two orchids of Kaiga forest (Western Ghats), India. J For Res 2012;23:453-60. [DOI]
23. Lubna, Asaf S, Hamayun M, Khan AL, Waqas M, Khan MA, Jan R, Lee IJ, Hussain A. Salt tolerance of Glycine max.L induced by endophytic fungus Aspergillus flavus CSH1, via regulating its endogenous hormones and antioxidative system. Plant Physiol Biochem 2018;128:13-23. [DOI]
24. Kalimuthu AK, Pavadai P, Panneerselvam T, Babkiewicz E, Pijanowska J, Mrówka P, Rajagopal G, Deepak V, Sundar K, Maszczyk P, et al. Cytotoxic potential of bioactive compounds from Aspergillus flavus, an endophytic fungus isolated from Cynodon dactylon, against breast cancer: Experimental and computational approach. Molecules 2022;27:8814. [DOI]
25. Gibson AM, Baranyi J, Pitt JI, Eyles MJ, Roberts TA. Predicting fungal growth: The effect of water activity on Aspergillus f lavus and related species. Int J Food Microbiol 1994;23:419-31. [DOI]
26. Gourama H, Bullerman LB. Aspergillus f lavus and Aspergillus parasiticus: Aflatoxigenic fungi of concern in foods and feeds: A review. J Food Prot 1995;58:1395-404. [DOI]
27. Choi MJ, Byun SH, Kim HS, Lee BM. Dietary exposure of aflatoxin B1 and cancer risk assessment. J Fd Hyg Safety 1995;10:81-7.
28. Maekawa N, Yamamoto M, Nishimura S, Kohmoto K, Kuwada K, Watanabe Y. Studies on host-specific AF-toxins produced by Alternaria alternata strawberry pathotype causing Alternaria black spot of strawberry. (1) Production of host-specific toxins and their biological activities. Ann Phytopathol Soc Jpn 1984;50:600-9. [DOI]
29. Bottini AT, Gilchrist DG. Phytotoxins. I. A 1-aminodimethylheptadecapentol from Alternaria alternata f. sp. lycopersici. Tetrahedron Lett 1981;22:2719-22. [DOI]
30. Headrick J, Pataky J. Maternal influence on the resistance of sweet corn lines to kernel infection by Fusarium moniliforme. Phytopathology 1991;81:268-74. [DOI]
31. Harper JK, Arif AM, Ford EJ, Strobel GA, Porco JAJ, Tomer DP, Oneill KM, Heider EM, Grant DM. Pestacin: A 1,3-dihydro isobenzofuran from Pestalotiopsis microspora possessing antioxidant and antimycotic activities. Tetrahedron 2003;59:2471-6. [DOI]
32. Strobel G, Yang X, Sears J, Kramer R, Sidhu RS, Hess WM. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology 1996;142:435-40. [DOI]
33. Grujić M, Dojnov B, Potočnik I, Atanasova L, Duduk B, Srebotnik E, Druzhinin IS, Kubicek CP, Vujčić Z. Superior cellulolytic activity of Trichoderma guizhouense on raw wheat straw. World J Microbiol Biotechnol 2019;35:194. [DOI]
34. Liu Q, Meng X, Li T, Raza W, Liu D, Shen Q. The growth promotion of peppers (Capsicum annuum L.) by Trichoderma guizhouense NJAU4742-based biological organic fertilizer: Possible role of increasing nutrient availabilities. Microorganisms 2020;8:1296. [DOI]