INTRODUCTION
Talaromyces is a member of the order Eurotiales within the family Trichocomaceae, recorded from soil, indoors, and food wastes, and has a worldwide distribution [1]. This fungal genus belongs to the phylum Ascomycota, the largest phylum of the kingdom fungi and comprises 64,000 fungal species [2]. The genus Talaromyces was introduced by Benjamin [3], and Talaromyces vermiculatus is the first species of this genus that produces soft, cottony-fruiting bodies (ascocarps) with cell walls made of tightly interwoven hyphae [4]. In addition, T. verruculosus, T. aculeatus, and T. apiculatus are the only species of Talaromyces that produce conidiophores with ampulliform phialides, which taper into very thin necks and have rough walled, globose conidia [2]. Talaromyces variabilis is synonymous with Penicillium. The P. variable is an anamorph species of fungus within the genus Penicillium, mainly isolated from permafrost deposits [5].
During the study and survey of fungal species from the field soil of Yangpyeong, South Korea, T. variabilis was also recovered. The aim of this study was to (i) investigate the T. variabilis isolate at the morphological and molecular level and (ii) to compare this T. variabilis isolate with previously reported T. variabilis isolate.
MATERIALS AND METHODS
Soil sample collection and fungal isolation
Soil samples were collected in 2017 from various locations in Yangpyeong, Korea. The GPS location of the soil sampling site was 37o28'52.81″ N and 127 °36'12.26″ E. Soil samples were taken from 0 to 15 cm depth after scraping and removing the surface leaf litter and other plant debris. Approximately 200-300 g of soil was collected from each collection site. The collected soil samples were dried at room temperature, ground, and sieved using an autoclave-sterilized brass sieve with an aperture size of 2 mm. Fungi were isolated using a serial dilution technique [6]. In detail, 1 g of soil sample was suspended in 9 mL of sterile distilled water and vigorously shaken for 3-5 min. The soil suspensions were then serially diluted in sterile distilled water. One milliliter of each 10-3, 10-4, 10-5, and 10-6 dilution was pipetted out and poured into Petri dishes containing potato dextrose agar (PDA; Difco Laboratories, Detroit, MI, USA), and the plates were incubated for 7 days at 25℃ until the growth of fungal colonies was observed. Morphologically distinct colonies were picked and purified by sub-culturing on plates containing PDA medium. Pure cultures of the fungal species were preserved in 20% glycerol stock at 4℃ for further studies.
Morphological characterization
Morphological study of KNU17-154 was performed based on growth characteristics on five different media: potato dextrose agar (PDA), oatmeal agar (OMA), Czapek yeast extract agar (CYEA), yeast extract sucrose agar (YESA), and malt extract agar (MEA). The strains were inoculated at three points on 9-cm petri dishes and incubated for 7 days at 25℃ in the dark. Media preparation was performed using the protocol described by Samson et al., [7]. After incubation, the colony diameter on PDA medium was measured. Colony color (front and back sides) and degree of sporulation were also observed. Colony colors were described using Kornerup and Wanscher [8]. Colony photographs of the fungal species were observed using a 3.1 CMOS digital camera (KOPTIC Korea Optics, Seoul, Korea) connected to an Olympus BX50F-3 microscope (Olympus Optical Co., LTD., Tokyo, Japan). Moreover, scanning electron microscope (SEM) images of the microstructures of the species were also observed using a LEO Model 1450VP Variable Pressure Scanning Electron Microscope (Carl Zeiss, Cambridge, Massachusetts, USA).
Genomic DNA extraction, PCR, sequencing, and phylogenetic analysis
Total genomic DNA of the isolates was extracted using the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. The internal transcribed spacer region (ITS) and calmodulin (CaM) gene were amplified using primers ITS1 (5'- TCCGTAGGTGAACCTGCG-3') and ITS4 (5'- TCCTCCGCTTATTGATATGC-3') [9] and CMD5 (5'- CCG AGT ACA AGG CCT TC-3') and CMD6 (5'- CCG ATA GAG GTC ATA ACG TGG-3'), respectively. The amplified PCR products were sequenced using an ABI Prism 3730 DNA analyzer (Applied Biosystems, Foster City, CA, USA). The conditions of the thermal cycles for the amplification and sequencing are presented in Tables 1 and Table 2. Reference ITS and CaM sequences were used to compare the sequences of GenBank at the National Center for Biotechnology Information (NCBI) using the basic local alignment search tool [10]. The nucleotide sequence was deposited in the culture collection of the National Institute of Biological Resources (NIBR, Incheon, Korea). Talaromyces variabilis (KNU17-154) was deposited at NIBR in 2017 under number NIBRFG0000501870. The nucleotide sequences of the study isolate were deposited in GenBank under the accession number MH231766. The sequences generated in this study were edited, merged, and proofread similar to those from GenBank using MEGA6 [11]. A neighbor-joining tree was constructed using the Kimura 2-parameter substitution model [12], and bootstrap analysis was performed with 1,000 replications to determine the support for each clade.

RESULTS
Scientific names: Talaromyces variabilis (Sopp) Samson, Yilmaz, Frisvad & Seifert, Studies in Mycology 70: 177 (2011) [MB#560676].
Synonymy: Penicillium variabile Sopp, Skrifter udgivne af Videnskabs-Selskabet i Christiania. Mathematisk-Naturvidenskabelig Klasse 11: 169 (1912) [MB#212359]
Colony morphology
The detailed morphology of the isolated colony, including the microstructures, is shown in Fig. 1. On PDA (potato dextrose agar), colonies grew slowly, attaining a diameter of 15-18 mm after 7 days at 25℃. The front and back sides of the colony were light orange in color, while the edges were creamy white (Figs. 1A and 1F). The sporulation was moderate to dense. The texture of the colony was floccose. The colony margins were irregular (Figs. 1A and 1B). On oatmeal agar (OMA), colonies grew fast, reaching a diameter of 25-30 mm after 7 days at 25℃ (Figs. 1B and 1G). The front side of the colony was green, and the back side of the colony was light yellow in color (Figs. 1B and 1G). The edges of the front side colony were white (Figs. 1A and 1G). The colony texture was floccose, and conidia were numerous. The sporulation was moderate to dense. The colony margins were irregular in shape. On Czapek yeast extract agar (CYEA), colonies grew slowly, attaining a diameter of 15-19 mm after 7 days at 25℃ (Fig. 1C and 1H). The front side of the colony was whitish orange and green, and the back side of the colony was dark yellow (Fig. 1C and 1H). The texture was floccose. Sporulation was dense, and the conidia were numerous. The colony margins were irregular. On yeast extract sucrose agar (YESA), the colony grew moderately, reaching a diameter of 25-30 mm after 7 days at 25℃ (Figs. 1D and 1I). The front side of the colony was light green to white, and the back side of the colony was light yellow (Fig. 1D and 1I). The texture was floccose, sporulation was moderate to dense, and the conidia were numerous. On malt extract agar (MEA), the colony grew slowly, attaining a diameter of 15-18 mm after 7 days at 25℃ (Fig. 1E and 1 J). The front and back sides of the colony were green and yellow in color, respectively (Fig. 1E and 1J). The margin was irregular and the texture was floccose. The sporulation was moderate to dense. Conidia were numerous.
Micro morphology
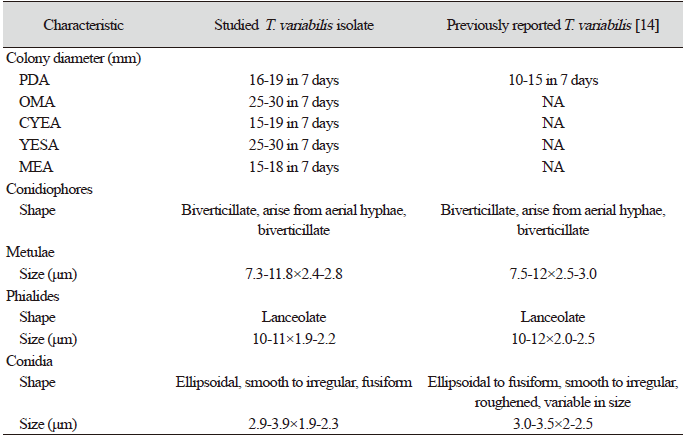
The detailed microstructures of the isolate are shown in Fig. 1. A brief description and comparison of the isolate is presented in Table 3. The conidiophores were biverticillate, arising from the aerial hyphae (Fig. 1l and 1M). Phialides are lanceolate with the size of [(10-11)×(1.9-2.2)] μm (Fig. 1L and 1M). Conidia were strongly ellipsoidal, smooth to irregular, and fusiform with a size of [(2.9-3.9)×(1.9-2.3) μm (Fig. 1K and 1N). Metulae were of [(7.3-11.8)×(2.4-2.8)] μm in size (Fig. 1L).
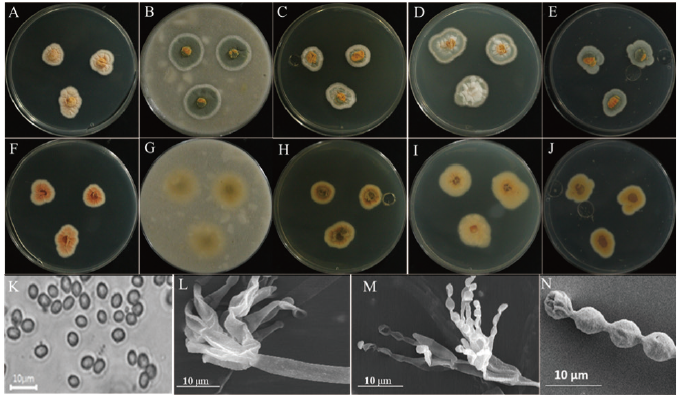
Fig. 1. Morphological characteristics of Talaromyces variabilis KNU17-154 grown on potato dextrose agar (PDA) at 25℃ for 7 days. Obverse colony from left to right (A-E) and reverse colony from left to right (F-J) grown on PDA, oatmeal agar (OMA), Czapek yeast extract agar (CYEA), yeast extract sucrose agar (YESA), and malt extract agar (MEA). Conidiophores and conidia produced on PDA (K and L) taken using a simple and scanning electronic microscope. Phialides observed in scanning electronic microscope (M) and scanning electronic microscope of conidia (N) produced on PDA.
Molecular phylogeny
Molecular analysis was performed to study the relationships among the isolate, KNU17-154, and the previously described reference isolate from NCBI BLAST. The regions for ITS sequence and CaM gene were used for molecular analysis. The results obtained from the gene sequence analysis of 18S-ITS1-5.8S-ITS2-28S rDNA sequences revealed that KNU17-154 was most closely related to Talaromyces variabilis (KF196915.1), creating a monophyletic clade group with 99% bootstrap value (Fig. 2). Moreover, for the precise identification of the isolate KNU17-154, CaM sequence data were obtained from the previously described T. variabilis isolate from the NCBI database. The neighbor-joining result suggests that our isolate is closely related to T. variabilis (KF196878.1), forming a monophyletic clade with a bootstrap value of 99 % (Fig. 3).

Fig. 2. Neighbor joining phylogenetic analysis of Talaromyces variabilis KNU17-154 and using the partial 18S-ITS1-5.8S-ITS2-28S rDNA region sequence obtained from field soil in Korea. Numerical values (>50) on branches represent the bootstrap values shown as the percentage of bootstrap replication from a 1,000-replicate analysis. ‘T’ indicates the type strain of the isolate.
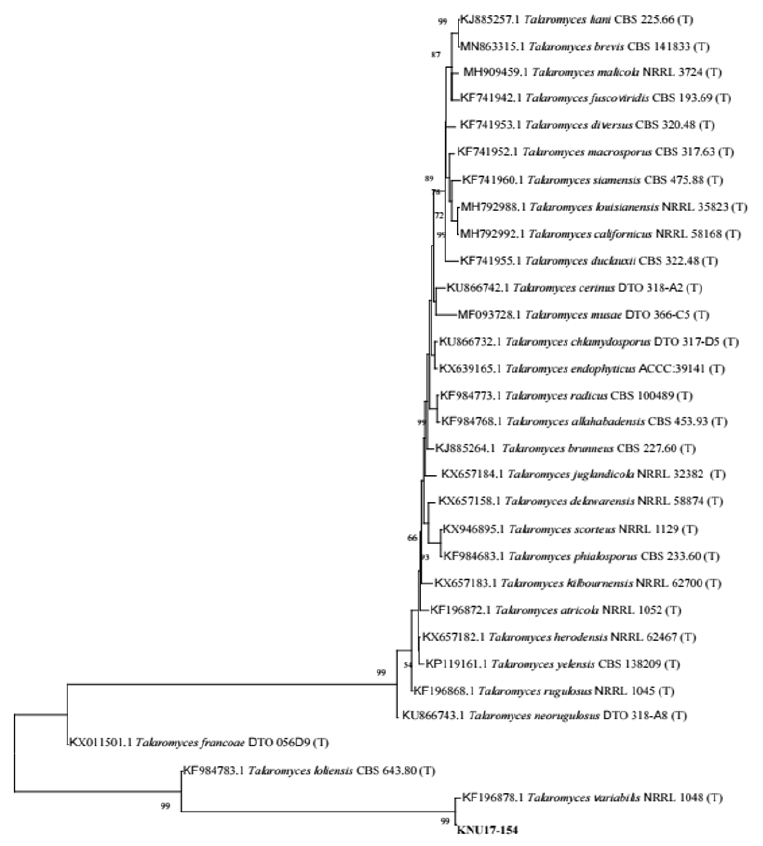
Fig. 3. Neighbor-joining phylogenetic analysis of Calmodulin (CaM) gene sequences of Talaromyces variabilis KNU17-154. Tree was constructed using MEGA6. Sequences obtained in the study are shown in boldface. Numerical values (>50) on branches are the bootstrap values as percentage of bootstrap replication from a 1,000 replicate analysis. Scale bar represents the number of substitutions per site.
DISCUSSION
During the fungal diversity survey in soils from various places in Korea, several fungal isolates were obtained through soil isolation and identification through morphological and molecular basis. Our study isolate KNU17-154, which was obtained from field soil in Yangpyeong, Korea, in 2017, has morphological characteristics similar to those of previously reported Talaromyces variabilis [13]. The shape, size, and structures of conidiophores, phialides, metulae, and conidia of KNU17-154 were found to be close, with only slight differences in size. DNA sequencing and neighbor-joining analyses are precise and reliable methods for fungal identification [14]. Similar to other fungi, T. convoluted the observation of small and large subunit ribosomal nucleic acids as well as ITS sequences [15]. However, consecutive analysis indicates that ribosomal genes are too conserved to differentiate the Talaromyces group of fungi [16]. In addition, ITS is currently authorized as the official DNA barcode for fungi [17]. In this study, we also used ITS sequences of our study to identify KNU17-154 isolate. The phylogenetic analysis according to ITS sequences results indicated that the fungal isolate KNU17-154 had 99% similarity with T.variabilis NRRL 1048 (Fig. 2). Moreover, ITS sequence variability can be used to differentiate species of various sections or series [17]; however, it is not always useful to distinguish species from the same section or series. To address this, multiple genetic markers including CaM gene were advocated to be used to identify fungal species accurately [18], and the sequencing of CaM gene can help identify Talaromyces species [15]. Following the above statement, we also analyzed the CaM gene sequences of KNU17-154. Phylogenetic analysis according to CaM result exhibited 99% similarity to KNU17-154 with T. variablis NRRL 1048 (Fig. 3). In conclusion, the fungal isolate KNU17-154 was identified as T. variabilis by morphological and molecular characterization. This fungus is important in the fields of microbiology and biotechnology, as it produces various secondary metabolites. This fungal isolate also has the potential to suppress damping-off of cucumbers and tomatoes [19]. Further studies regarding the biocontrol activities of this untapped fungal isolate might be of interest to researchers in the future.
ACKNOWLEDGEMENTS
This study was financed by the Ministry of Environment (MOE) of the Republic of Korea from the National Institute of Biological Resources (NIBR) grant (NIBR2014-01205) on the survey and discovery of indigenous fungal species in Korea. This study has been worked with the support of a research grant of Kangwon National University in 2019.

