Introduction
Adzuki bean (Vigna angularis L.), belonging to the family Fabaceae, is an annual climber referred to as azuki bean, aduki bean, atsumi bean, red bean, or red mung bean [1]. It is widely cultivated throughout East Asia and is commercially produced in the United States, South America, India, New Zealand, Congo, and Angola [2]. Adzuki bean is grown and used mainly for edible purposes. It has become an important economic crop in East Asia, mainly in the cultivated areas of China (670,000 ha), Japan (60,000 ha), South Korea (25,000 ha), and Taiwan (15,000 ha) [1]. Adzuki bean seeds contain essential plant fatty acids such as linoleic acid, palmitic acid, linolenic, etc., and several phytochemicals, including phytates and polyphenols [3]. In Korea, the adzuki bean is the second most important legume crop after soybean. It is used in traditional Korean dishes such as adzuki porridge, adzuki rice cake, adzuki noodles, adzuki jelly, and shaved ice with adzuki bean [4].
Diseases related to the adzuki bean have been reported worldwide, such as powdery mildew caused by Erysiphe spp., brown stem rot caused by Phialophora gregata, root rot caused by Fusarium spp., bacterial blight caused by Xanthomonas campestris, and viral diseases such as adzuki bean mosaic potyvirus (AzMV), blackeye cowpea mosaic potyvirus (BICMV), bean common mosaic potyvirus (BCMV), bean yellow mosaic potyvirus (BYMV), cucumber mosaic cucumovirus (CMV), and alfalfa mosaic virus (AMV) [5-7]. Among them, powdery mildew (Podosphera phaseoli), circular leaf spot (Ascochyta phaseolorum), leaf spot (Cercospora cruenta), anthracnose (Colletotrichum spp.), and damping off (Fusarium sp.) are known as fungal diseases that occur on adzuki beans in Korea [8]. Fusarium root rot of adzuki bean was first observed in 1996-1998 in eastern Hokkaido, Japan [9]. Outbreaks of adzuki bean root rot disease have been continuously reported in several countries, including the United States and China, etc. However, according to the 6th edition of the List of Plant Diseases in Korea, adzuki bean root rot caused by Fusarium species has not been reported in Korea.
F. oxysporum is known as the Fusarium species that causes root rot on adzuki beans in China [10] and Japan [11], while F. falciforme has been identified as the causive agent of adzuki bean root rot in Brazil [12]. In 2012, F. azukicola [9] was additionally reported as a new species in Japan. When these Fusarium species cause root rot in adzuki beans, it leads to stunting, leaf yellowing, vascular discoloration, and wilting [10,11].
In this study, we reported Fusarium root rot of adzuki beans by isolating Fusarium species, causing disease in the adzuki bean fields of Korea. Species were identified by analysis of morphological, cultural, and molecular characteristics and the pathogenicity of the isolates was confirmed.
Materials and Methods
Sampling and fungal isolation
In July 2020, samples showing wilting symptoms were collected from five adzuki bean fields in Yeosu, Korea. To isolate the fungal isolates, the discolored tissues inside the root and stem of the diseased plants were cut and surface-sterilized with 1% sodium hypochlorite (NaOCl) for 1 min followed by rinsing twice in sterile distilled water. The tissue pieces were placed on water agar (WA) and incubated at 25℃ for 5 days. The colonies growing on the sample pieces were observed under a microscope. Fusarium isolates were isolated by the single spore isolation method [13]. All isolates were cultured in potato dextrose agar (PDA) slants and stored at 10℃ until further use.
Investigation of morphological and cultural characteristics
To investigate the morphological characteristics, three isolates were grown on PDA and carnation leaf agar (CLA) [14]. For the morphological identification, isolates were cultured on CLA at 25℃ under near ultraviolet (NUV)/dark (12 hrs/12 hrs) conditions for 14 days. The characteristics of macro- and micro-conidia, the presence of chlamydospores, the morphology of phialides, and sporodochial formation were investigated. The colony color, presence of aerial mycelium, pigmentation into agar, and growth rate were investigated on PDA at 25℃ with the dark condition for 7 days.
DNA extraction and molecular identification
Genomic DNA was extracted from the mycelia powder using a Maxwell® RSC PureFood GMO and Authentication Kit (Promega, Madison, WI, USA) according to the manufacturer’s instructions. Each isolate was cultured in 20 mL of potato dextrose broth (Difco, Bergen, USA), at 25℃ for 7 days. The cultured mycelia were harvested using miracloth and freeze-dried overnight. The dried mycelia were ground using a homogenizer. The mycelial powder was vortexed with 20 μL of RNase A and 30 μL of proteinase K, and incubated at 65℃ for 30 min in a heating block. Samples were centrifuged at 14,000 rpm for 5 min, then 400 μL of the supernatant was recovered. From this supernatant, genomic DNA was finally extracted using Maxwell® RSC. All genomic DNA was stored at −20℃ before use. For the molecular identification of the isolates, the translation elongation factor 1 alpha (TEF) and RNA polymerase Ⅱ second largest subunit (RPB2) coding regions were selected [15] and amplified using the primers EF1/EF2 and fRPB2-5f2/fRPB2-11aR [16], respectively. PCR amplification was performed using a 50-μL mixture containing 3-μL DNA templates (33 ng/μL), 3 μL of each F/R primer (5 pmoles/μL), 5 μL of 10× nTaq-Tenuto buffer (nTaq-Tenuto (Mg2+ plus), Korea), 5 μL of 2-mM dNTP mixture, 30 μL of sterile water, and 0.5 μL of nTaq-Tenuto polymerase. The PCR conditions for DNA amplification of the TEF included initial denaturation at 95℃ for 4 mins; followed by 34 cycles of denaturation at 95℃ for 30 s, annealing at 58℃ for 45 s, extension at 72℃ for 50 s; and final extension at 72℃ for 7 min [17]. For RPB2, the PCR conditions were 95℃ for 2 min; followed by 34 cycles of 95℃ for 30 s, 56℃ for 30 s, 72℃ for 2 min; and final extension at 72℃ for 5 min [18]. All final products of PCR were observed by electrophoresis on a 1.4% agarose gel. The PCR products were purified using a Wizard® SV Gel and PCR Clean-up System Kit (Promega, San Luis Obispo, CA, USA) according to the manufacturer’s instructions. PCR products were sequenced in both directions by Bionics Co., Ltd. (Seoul, Korea). Sequences of the isolates and reference strains obtained from the National Centre for Biotechnology Information (NCBI) GenBank were aligned using the MUSCLE method in the MEGA-X program [19,20]. Phylogenetic trees were constructed based on the neighbor-joining(NJ) and Kimura 2-parameter models and verified by 1,000 bootstrap replicates [21]. Fusarium solani NRRL 25388 was used as an outgroup.
Pathogenicity test
The pathogenicity test of the isolates was carried out on adzuki bean (cv. Arari) using the soil inoculation method [22]. To prepare the soil inoculum, 240 g of dry sand, 26 g of cornmeal, and 65 mL of distilled water were mixed in a 500-mL Erlenmeyer flasks. The mixtures were autoclaved twice at 121℃ for 30 min and inoculated by adding 15 PDA disks (5-mm) containing isolate mycelia. Pure PDA disks were used as controls. Samples were incubated at 25℃ for four weeks without shaking. Four weeks after inoculation, the cornmeal sand inoculum and sterilized soil were mixed at a volume ratio of 3:7 and divided into 200 mL for each pot. Two germinated seeds were planted in each pot, and three replicates were performed. All the plants were grown in the plant growth room at 25-27℃, with a photoperiod of 12 hr per day. Three weeks after sowing, disease incidence was scored using a 0-4 point scale (Fig. 1).
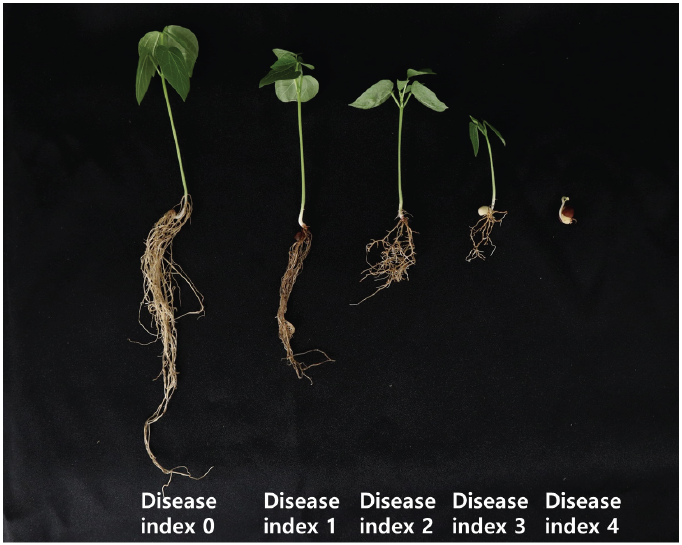
Fig. 1. Disease index of Fusarium root rot in adzuki bean plants. 0=no symptoms; 1=necrosis of root tissue and less than 30% of average root volume; 2=necrosis of root tissue and less than 31-60% of average root volume; 3=root tissue necrosis, lack of root volume more than 61% and poor growth; 4=complete necrosis of root tissue and no roots or plants death.
Additionally, to determine whether these isolates could cause bakanae disease in rice plants, artificial inoculation of rice seeds was performed using the seed soaking method [23]. Disinfected rice seeds (cv. Sindongjin) were put into the spore suspension at a concentration of 106 spores/mL and incubated at 25℃ for two days. The controls were soaked in sterilized distilled water. Three rice seeds were sowed in a test tube (200 mm×30 mm) containing 10 mL of 0.6% WA medium (replicated five times) and incubated at 30℃ on 12-hr light/ 12-hr dark cycle. After 2 weeks, disease incidence was scored using a 0–5 point scale [23], where 0=symptomless, 1=plants pale green, 2=plants yellow-green and slightly elongated compared to control, 3=plants yellow-green, slightly elongated compared to scale 2, 4=plants yellow, and twice as elongated compared to control, 5=plants with severe bakanae symptoms or plant death caused by bakanae. The score obtained was standardized on a 0-100 scale.
Results
Symptoms and fungal isolation
A total of five adzuki bean fields in Yeosu were investigated. Diseased plants were observed in two places. The incidence of wilting symptoms was less than 1% in the entire field, with symptoms such as yellowing, stunting, wilting, and browning of the roots and stems (Fig. 2). Three Fusarium isolates were obtained from the infected adzuki beans.

Morphological and cultural characteristics
The morphological and cultural characteristics of the three isolates, including the growth rate, aerial mycelia, colony texture, macroconidia, microconidia, etc., were similar among the three isolates (Fig. 3). The growth rates on PDA were approximately 9-11 mm per day (diameter). The color of the aerial mycelium was white to violet, the colony texture was fluffy, and the pigmentation into the agar was dark magenta, yellow, grayish violet, or pale violet. Sporodochia was observed as pale orange to orange color on pieces of carnation leaf in CLA. The shape of macroconidia was slender with no significant curvature, blunt, apical cells, and poorly developed foot-shaped basal cells. The size was 29.3–55.6×2.3–4.2 µm, and the number of septa was 3-5 septate. In the case of microconidia, the shape was obovoid, oval, and clavate, the size was 5.8–9.4×2.3–3.9 µm, and the number of septa was 0-1. The microconidia were formed into chains, and chlamydospores were not observed. Details on the morphological and cultural characteristics of the previously known F. fujikuroi and the isolate used in this study were described in Table 1.

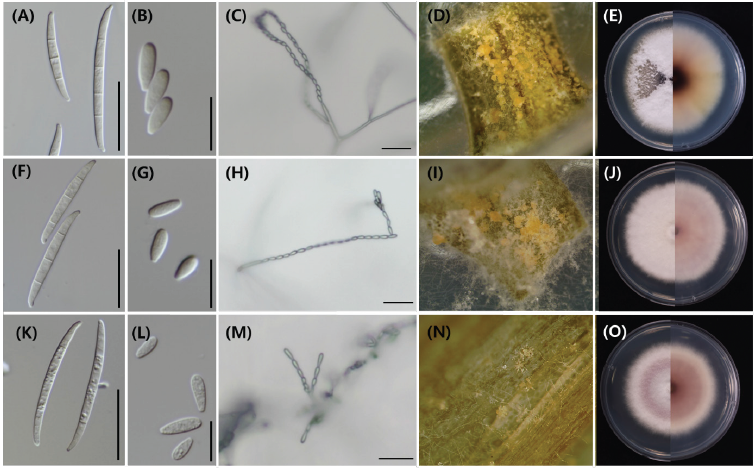
Fig. 3. Morphological and cultural characteristics of Fusarium fujikuroi isolates collected from adzuki beans. A, F, K, Macroconidia. B, G, L, Microconidia. C, H, M, Microconidia in chains. D, I, N, Sporodochia on carnation leaf agar (CLA). E, J, O, Colony culture on potato dextrose agar (PDA); A-E, NC20-737. F-J, NC20-738. K-O, NC20-739. Scale bars of macroconidia, microconidia, and chain are 25, 10, and 25 μm, respectively.
Molecular identification
The PCR amplification sizes of RPB2 and TEF were approximately 1,800 bp and 600-800 bp, respectively. In phylogenetic analysis, three isolates were grouped into one clade with reference strains of Fusarium fujikuroi NRRL 13566 and NRRL 5538 obtained from NCBI GenBank (Fig. 4). For the RPB2 gene, the three isolates were 100% identical to the reference strains, NRRL 13566 and NRRL 5538. For the TEF gene, the isolates were 100% similar to NRRL 5538 and 98% identical to NRRL 13566. The sequences of the RPB2 and TEF regions of NC20-737, NC20-738, and NC20-739 were deposited in GenBank with the accession numbers ON638992, ON548181; ON638993, ON548182; ON638994, ON548183, respectively.
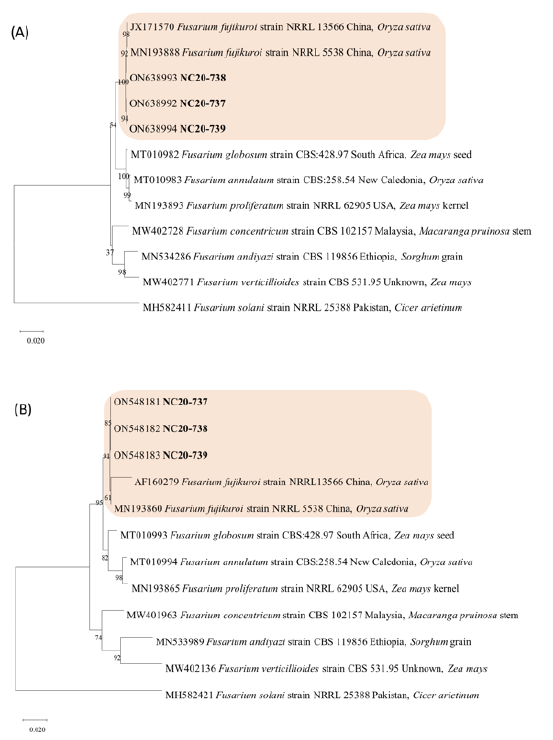
Fig. 4. Phylogenetic trees of F. fujikuroi obtained from wilted adzuki bean plants in Korea. The trees were constructed with (A) RNA polymerase Ⅱ second largest subunit (RPB2) and (B) translation elongation factor 1 alpha (TEF) gene nucleotide sequences. The number in each branch indicates bootstrap values obtained after a bootstrap test with 1,000 replications. The scale bar represents 0.02 nucleotide substitutions per site.
Pathogenicity test
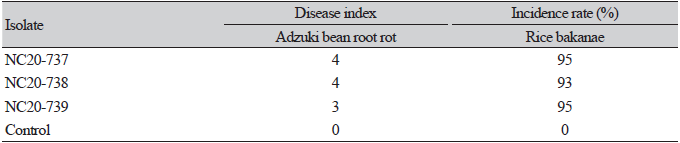
Three weeks after inoculation, the adzuki bean plants had few roots and they showed browning, rot, stunted growth, or death. Control plants did not show any symptoms. After the pathogenicity test, F. fujikuroi was reisolated from inside the root and stem tissues of the inoculated plants; therefore, Koch’s postulates were fulfilled. The results of the pathogenicity test with identical isolates for bakanae of rice, rice seedlings showed typical symptoms of bakanae, such as plant yellowing and abnormally elongated height (Table 2).
Discussion
This study confirmed that Fusarium fujikuroi is the causal agent of the root rot and wilt of adzuki beans in Korea. Throughout the world, various Fusarium species have been suggested as pathogens of legumes. Among them, F. fujikuroi is commonly known to cause rice bakanae and Fusarium wilt in soybeans and is known to cause wilt disease in adzuki beans by artificial inoculation [24,25]. In this study, F. fujikuroi, isolated from the adzuki bean cultivation fields caused wilting and root rot symptoms. Although Fusarium spp. have been reported to cause damping-off, the symptoms of damping-off mainly appear near the upper part of the soil, whereas root rot is characterized by gradual wilting symptoms after infecting the roots.
Fusarium species are important plant pathogens that affect diverse hosts, but identification at the species level is complex [26,27]. Therefore, we used the morphological features and molecular markers for Fusarium species identification in this study. Among the morphological characteristics, the Fusarium fujikuroi species complex (FFSC) [28] formed microconidia in chains along with macroconidia and sporodochia, as reported previously for F. fujikuroi species [27,29]. These characteristics were unique to F. fujikuroi. Although all three isolates were from the same species, there were slight differences in the pigmentation on PDA, macroconidia width, and amount of sporodochia formation. Leslie et al. [27] indicated that many isolates of F. fujikuroi form very few or no sporodochia and show pigmentation ranging from no pigmentation to grayish orange, violet gray, dark violet, or dark magenta. Due to intraspecific variation and morphological similarities among Fusarium species, it is impossible to identify species based on morphological characteristics alone.
Phylogenetic analysis to distinguish closely related species within the FFSC, such as F. fujikuroi, F. verticillioides, and F. proliferatum, has been conducted previously using protein coding genes, including TEF, RPB2, beta-tubulin, calmodulin, and histone genes in many previous reports [27,30,31]. Among these genes, the TEF and RPB2 regions are helpful for Fusarium species identification because they meet three important criteria for phylogenetic species identification [15]. Therefore, we used the TEF and RPB2 genes, and the phylogenetic tree was performed in three cases: 1) TEF, 2) RPB2, and 3) TEF+RPB2. In all three cases, isolates were clustered in the same clade as the F. fujikuroi representative isolates NRRL 13566 and NRRL 5538 and differentiated from other species belonging to FFSC (Fig. 4). From these results, it is thought that identification at the species level is possible from sequencing of the TEF or RPB2 region alone.
In the pathogenicity test results, F. fujikuroi was able to infect adzuki bean roots. The inoculated adzuki beans grew slowly from the beginning of growth or the seeds rotted underground. After three weeks, the plant height and leaf size were smaller than those of control plants. When these plants were uprooted, it was confirmed that the root amount was very small compared to the control, and the roots were brown. According to Aoki et al. (2012) [9] F. azukicola caused root rot and reduced plant height and dry weight in adzuki beans, consistent with the results of this study. In this pathogenicity test, Koch’s postulates were fulfilled, and the symptoms of inoculated plants were similar to typical symptoms described in other Fusarium root rot/wilt reports [9,10,32,33]. To the best of our knowledge, this is the first report of Fusarium root rot of adzuki bean caused by F. fujikuroi worldwide.
F. fujikuroi are known to cause bakanae disease in rice (Oryza sativa L.), characterized by the formation of elongated stems by the rice plant in response to gibberellins [34]. Therefore, we conducted an additional pathogenicity test on rice to determine whether F. fujikuroi collected from adzuki beans could also cause rice bakanae disease. Plants infected with F. fujikuroi in the current study showed the same symptoms as those of rice bakanae disease caused by F. fujikuroi in a previous study [21], with a high incidence rate. Therefore, it was proved that the isolates of this study were pathogenic not only to adzuki beans but also to rice.
In this study, we confirmed that it is challenging to identify Fusarium species only through morphological features alone. We also demonstrated that F. fujikuroi causes root rot in adzuki beans and bakanae disease in rice. It is necessary to analyze the genetic and pathogenic differences in F. fujikuroi isolated from various hosts such as rice, adzuki bean, and soybean, and to study the host-pathogen interaction. In addition, the isolates could be used for breeding adzuki bean root rot-resistant cultivars.

